
Actin type and distribution in erythrocytes
Abstract
Erythrocytes transport oxygen from the lungs to the tissues. The excess surface area together with the elasticity of the erythrocyte cell membrane provides the flexibility needed to pass through the microvasculature where the oxygen exchange occurs. Although the architecture of the red cell and its membrane-associated cytoskeletal network is known in general, the factors that control the characteristic shape change during echinocyte formation are poorly understood. In this short report we show that in echinocytes a completely reorganized membrane cytoskeleton with a box-like structure of actin filaments prevailed indicating the importance of the actin cytoskeleton during echinocyte formation.
The red blood cell membrane is an elastic shell with remarkable properties. Peripheral erythrocytes are cell remnants, since they have lost their nuclei, cytoplasmic organelles and are no longer able to synthesize proteins and to proliferate. However, they are absolutely essential for the survival of any macro organism with a blood circulation, because they transport the majority of the necessary oxygen to the cells of the body. Human erythrocytes circulate in the body for about 120 days and are normally discocytes with a biconcave shape and a diameter of ∼7.5 μm, a surface area of ∼140 μm2 and a volume of ∼100 μm3 [1]. The excess surface area of erythrocytes (allowing the flattened shape under resting conditions), together with the elasticity of their membranes, provides them with the flexibility needed to pass through very small capillaries with diameters as small as 2–3 μm [2, 3]). The erythrocyte membrane consists of a phospholipid bilayer with embedded membrane proteins and an underlying membrane cytoskeleton.
Although the architecture of the red cell and its membrane-associated cytoskeletal network is known in general [4], the factors that determine and control the characteristic biconcave shape and the shape change during echinocyte formation are poorly understood [5, 6]. Figure 1 shows erythrocytes in autologous plasma with their characteristic biconcave discoidal shape – called discocytes - on a glass cover slip.
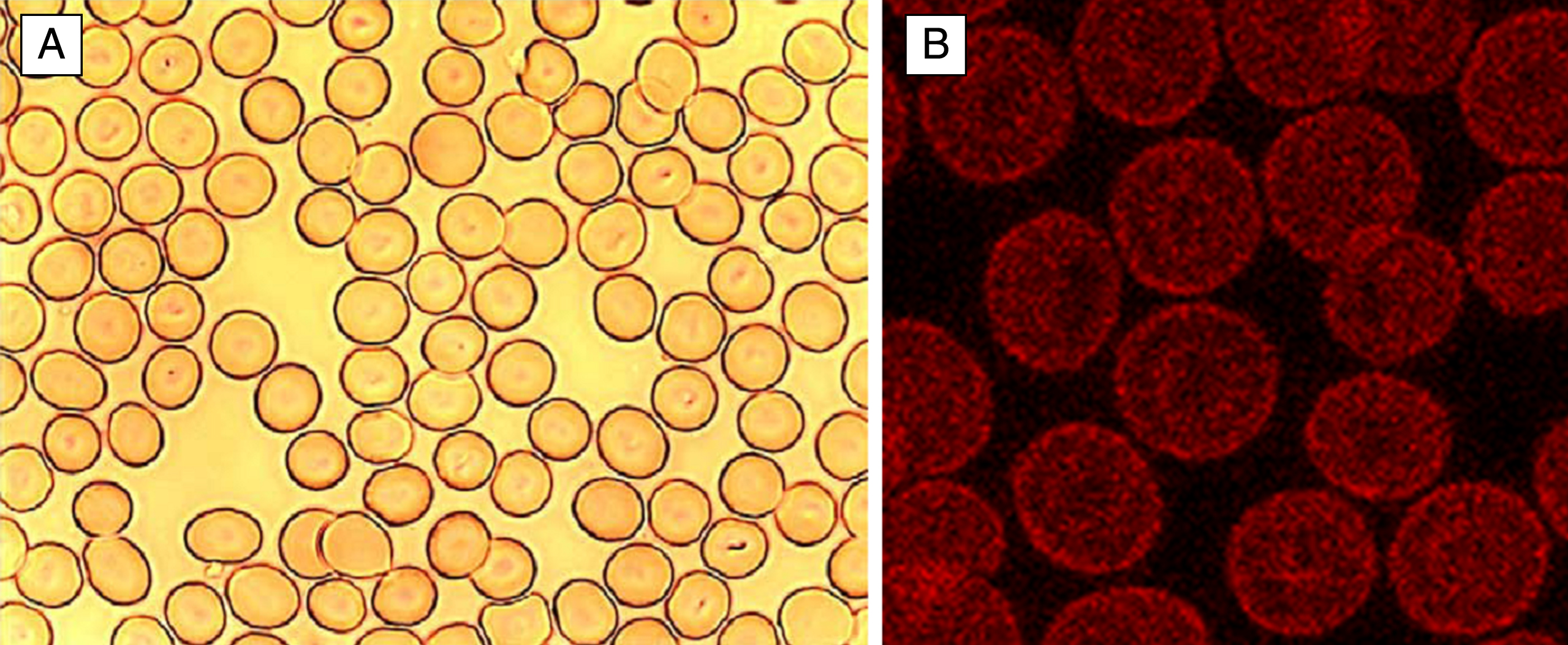
Fig.1
Normal human discocytes on a glass cover slip in autologous plasma. A: light microscopy, B: Phalloidin-rhodamine stained actin in discocytes visualized with a confocal laser scanning microscope.
Changes of erythrocyte morphology are affected by their membrane cytoskeleton. Actin is a key component of the membrane cytoskeleton [7], which links together with other elements the membrane cytoskeleton to the erythrocyte cell membrane. Actin of erythrocytes can be stained either with antibodies against β-actin (staining globular G-actin and fibrillar F-actin) or with phalloidin-rhodamine (staining F-actin oligomers described to consist of at least 16 monomers). β-actin stain resulted in globular red dots with a certain enhancement at the cell periphery, while phalloidin-rhodamine stain resulted in more homogenously stained erythrocytes, again with a certain enhancement at the periphery in a lot of cells [8].
The remarkable elasticity of the erythrocyte membrane is achieved by the combination of two properties: i) the elastic lipid/protein bilayer of the plasma membrane and ii) the quasi two-dimensional macro-molecular actin-spectrin network, which is loosely coupled to the bilayer.
In discocytes, the membrane cytoskeleton is displayed in a well-rounded shape consistent with the shape of discocytes suspended in autologous plasma (Fig. 1A). The Phalloidin-Rhodamine stained actin component coloured in red was inhomogeneously distributed over the cells. Thus, stained erythrocytes clearly revealed a more or less globular arrangement of actin and in most cells concentrical rings of globular actin, one ring at the periphery and another one around the centre of the erythrocytes were visible (Fig. 1B).
Echinocytic transformation of erythrocytes was assumed to occur when foreign molecules are preferentially intercalated in the outer hemi-leaflet of the red blood cell membrane. This was hypothesized to lead to an expansion of the outer cell membrane hemi-leaflet relative to the inner hemi-leaflet, following the bilayer couple theory [9]. Such an intercalation in the outer hemi-leaflet could result also in a formation of spicules. The precise interactions leading to the echinocyte formation are currently unknown. Figure 2B shows that the formation of echinocytes is associated with a complete deformation of the membrane cytoskeleton. The staining of the membrane cytoskeletal actin component showed in most cells a box-like arrangement of actin at the rim of the echinocytes. These findings were completely different from those found in discocytes and underline the importance of the membrane cytoskeleton for the shape of erythrocytes [10].
Fig.2
Echinocytes on a glass cover slip in autologous plasma. A: light microscopy, B: actin stained (phalloidin-rhodamine) erythrocytes visualized with laser scanning microscope.
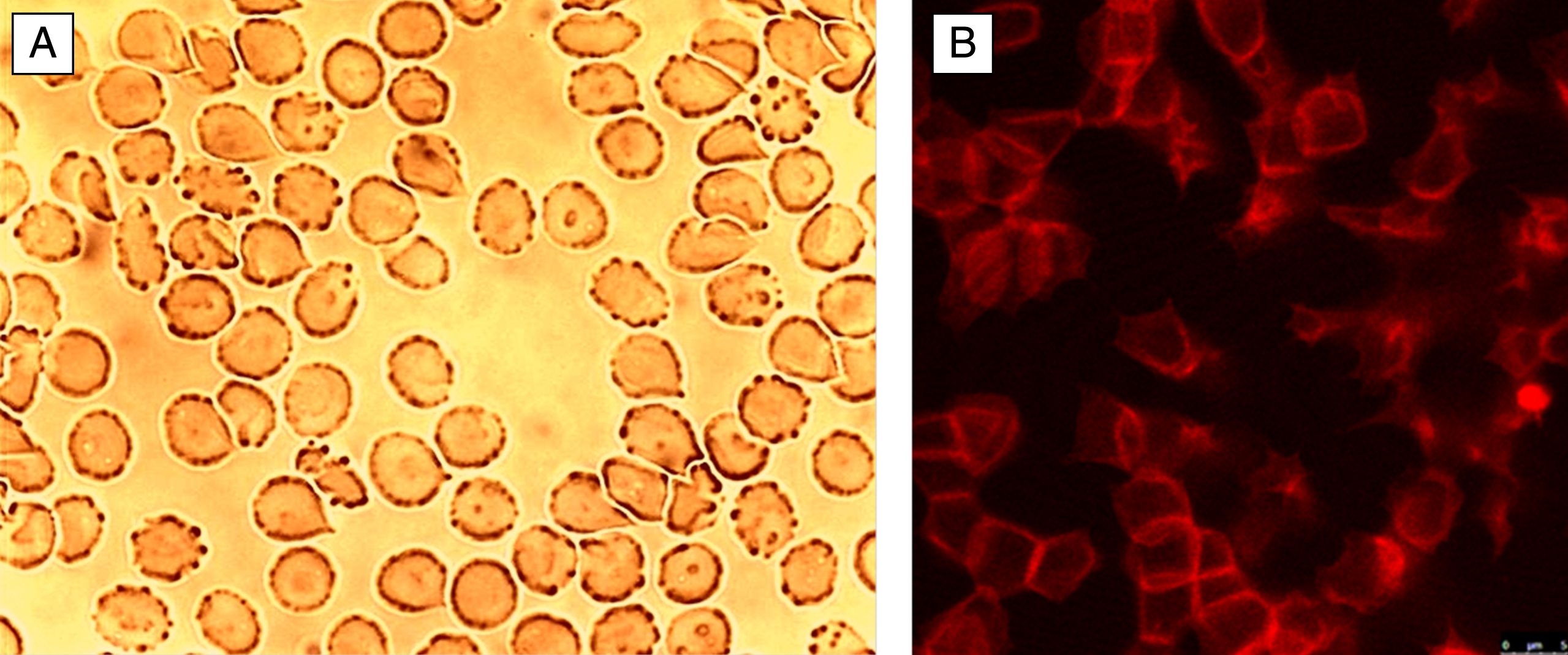
This reorganization of the cytoskeleton is not visible using a light microscope, here only the membrane with spicules are visible which mark the echinocyte formation.
Conclusion
While in human discocytes a more or less homogenous distribution of fine grained globular actin in the membrane cytoskeleton could be demonstrated, in echinocytes a completely reorganized membrane cytoskeleton with a box-like structure of actin filaments prevailed indicating the importance of the actin cytoskeleton during the echinocyte formation.
References
[1] | Evans E , Fung YC . Improved measurement of the erythrocyte geometry. Microvasc Res. (1972) ;4: :335–47. |
[2] | Hauw JJ , Berger B , Escourolle R . Ultrastructural observations on human cerebral capillaries in organ culture. Cell Tissue Res. (1975) ;163: :133–50. |
[3] | Kaliviotis E . Mechanics of the red blood cell network. Journal of Cellular Biotechnology. (2015) ;1: :37–43. |
[4] | Bennett V , Baines AJ . Spectrin and ankyrin-based pathways: Metazoan inventions for integrating cells into tissues. Physiol Rev. (2001) ;81: :1353–92. |
[5] | Diez-Silva M , Dao M , Han J , Lim C-T , Suresh S . Shape and Biomechanical Characteristics of Human Red Blood Cells in Health and Disease. MRS Bull. (2010) ;35: :382–8. |
[6] | Reinhart WH . Peculiar red cell shapes: Fåhraeus Lecture 2011. Clin Hemorheol Microcirc. (2011) ;49: :11–27. |
[7] | Matsuoka Y , Li X , Bennett V . Adducin is an in vivo substrate for protein kinase C: Phosphorylation in the MARCKS-related domain inhibits activity in promoting spectrin-actin complexes and occurs in many cells, including dendritic spines of neurons. J Cell Biol. (1998) ;142: :485–97. |
[8] | Franke RP , Scharnweber T , Fuhrmann R , Krüger A , Wenzel F , Mrowietz C , Jung F . Distribution of actin of the human erythrocyte membrane cytoskeleton after interaction with radiographic contrast media. Clin Hemorheol Microcirc. (2013) ;55: (4):481–90. |
[9] | Sheetz MP , Singer SJ . Equilibrium and kinetic effects of drugs on the shape of human erythrocytes. J Cell Biol. (1976) ;70: :247–51. |
[10] | Franke R-P , Krüger A , Scharnweber T , Wenzel F , Jung F . Effects of Radiographic Contrast Media on the Micromorphology of the Junctional Complex of Erythrocytes Visualized by Immunocytology. Int J Mol Sci. (2014) ;15: :16134–52. |


