
Respiratory virus infections and adenovirus characteristics during acute exacerbation of chronic obstructive pulmonary disease
Abstract
BACKGROUND:
Chronic obstructive pulmonary disease (COPD) is a common respiratory disease globally, characterized by obstructive ventilatory disorder under pulmonary function tests. Recent years have witnessed a yearly increase in the prevalence of COPD.
OBJECTIVE:
To investigate the impact of respiratory virus infections on patients with acute exacerbation of chronic obstructive pulmonary disease (AECOPD), and to perform sequencing typing and mutation analysis of viruses with high detection rate.
METHODS:
A total of 1523 inpatients with AECOPD admitted to our hospital from April 1,2020 to March 30,2022 were collected and divided into two groups: the infected group (
RESULTS:
In the infected group, the proportion of patients with palpitations (49.44% VS 8.07%,
CONCLUSION:
Respiratory viruses are common in patients with poor prognosis of AECOPD, especially adenovirus, respiratory syncytial virus. Respiratory virus infections will lead to the deterioration of patients with AECOPD, accompanied by longer treatment cycles and poor prognosis.
1.Introduction
Chronic obstructive pulmonary disease (COPD) is a common respiratory disease globally, characterized by obstructive ventilatory disorder under pulmonary function tests. Recent years have witnessed a yearly increase in the prevalence of COPD [1]. COPD, if progressed without timely control, may cause changes in the physiological structure of bronchi, alveoli and pulmonary vessels, resulting in imbalance of V/Q ratio and dysfunction of pulmonary ventilation [2]. When COPD is stimulated by physical and chemical factors or infected by bacteria and viruses in the stable period, the disease symptoms are more serious than usual, or the expectoration amount of sputum increases or the color changes. In such cases, the oral or inhaled medications used to maintain stabilization are no longer effective in controlling symptoms, and COPD turns into acute exacerbation of COPD (AECOPD), leading to a dramatic increase in the rates of disability and death.
As far as the deterioration of AECOPD itself is concerned, it is a comprehensive process of multiple factors such as individual patients, respiratory pathogen infection and environment [3]. Respiratory virus infection, as one of many pathogenic factors, is regarded as the primary trigger of AECOPD. However, no clear explanation of the mechanism leading to respiratory exacerbation has been presented. It is now generally accepted that the main mechanism of COPD deterioration is that respiratory virus colonizes alveolar epithelial cells, causing cell damage and shedding, promoting the release of inflammatory factors and so on, resulting in cell hyperresponsiveness and airway remodeling leading to tracheal stenosis. In the mouse model of chronic obstructive pulmonary disease, the infection with respiratory syncytial virus can obviously aggravate the symptoms of mice, and increase the expression of leukemia inhibitory factor (LIF) in the lungs. thus enhancing lung injury and airway hypersensitivity; In contrast, LIF and its receptor are instead reduced in the lungs of cigarette-smoking animal models. However, after comparing LIF factors in alveolar lavage fluid (BALF) from smokers, COPD patients and normal population, opposite conclusions were reached [4]. Another study on COPD and congestive heart failure (CHF) concluded the same findings, i.e. respiratory syncytial virus can cause acute exacerbation of this type of disease. The author also innovatively put forward that active immunization with monoclonal antibodies can benefit such patients more [5]. Human rhinovirus can also cause acute exacerbation of COPD (AECOPD) and acute attack of asthma, and can cause lung inflammation continuously after being infected with rhinovirus, leading to the accumulation of macrophages and CD8T cell subsets and the progress of emphysema [6]. In 1998, the Italian team M Saetta proved for the first time that persistent latent virus infection can induce a series of actions in CD8+ cells to remodel the airways, thus continuously worsening COPD [7]. Latent virus infections in COPD are dominated by herpesvirus and adenovirus, where adenoviral infections that occur during childhood when the body is immunocompromised will lead to inflammatory reactions such as bronchiolitis. These inflammatory processes lead to airway growth malformations that promote COPD disease to be more likely to occur in adulthood. After treatment and intervention, adenovirus will lurk in respiratory epithelial cells, and the E1A gene of the latent adenovirus will be activated after inhaling particulate pollutants from cigarettes or air. As a transcription activator, E1A gene will cooperate with the inflammatory reaction of human alveolar epithelial cells to cigarette smoke, resulting in a large amount of mucus secretion blocking the airway [8]. More studies have shown that when stimulated by other respiratory viruses, latent adenovirus will synergize with other respiratory viruses to produce higher levels of interleukin-8 and ICAM-1, which will aggravate the pulmonary inflammatory response [9].
Considering that virus infections are seasonal, different viruses have different seasons of high prevalence [10, 11, 12, 13]. Meanwhile, virus infections are also geographically differentiated, with prevalence rates varying from region to region. In the reports of western countries, picornaviruses are the main ones, and influenza viruses are the main ones in Asia [14]. In the present study, patients with AECOPD were screened for five common respiratory viruses to analyze the impact of virus infections on treatment prognosis, and adenoviruses with high detection rates were genetically characterized. In doing so, we aimed to lay a solid theoretical foundation for early screening and interventional treatment of respiratory viral infections in the clinical setting, and provide data support for the prevention and control of respiratory viral infections in those with AECOPD.
2.Study subjects and methods
2.1Study subjects and methods
We collected 1523 inpatients diagnosed with AECOPD in the emergency department, respiratory and critical care department and critical care department of our hospital from April 1, 2020 to March 30, 2022 by convenience sampling method, and divided them into two groups based on the presence or absence of virus infections: the infected group (532) and the non-infected group (991 cases). The inclusion criteria were as follows: (1) patients over 18 years old; (2) thoSe who met the diagnostic criteria of AECOPD in 2017 COPD Global Strategy Guidelines [15]; and (3) those themselves or family members consented to sampling and sent the samples for testing. The exclusion criteria were as follows: (1) patients with immune hypoimmunity or immune disorders such as rheumatoid arthritis, dermatomyositis, systemic lupus erythematosus and Sjogren’s syndrome; (2) those with immune system dysfunction such as AIDS; (3) those with negative pharyngeal swab testing for the human endogenous genes; and (4) those with incomplete clinical data. This study was approved by the Ethics Committee of our hospital. All patients or their families signed an informed consent form.
2.2Study methods
2.2.1Collection of throat swab samples
All the included samples were smeared by professionally trained medical staff on the throat wall and tonsils of the patients when they were admitted to the hospital. Immediately after the sampling, the virus throat swab tube was stored in the refrigerator at
2.2.2Detection of respiratory virus
(1) Sample treatment: Firstly, the throat swab tube is repeatedly shaken upside down, so that the respiratory virus and human throat mucosa cells attached to the cotton swab are evenly mixed with the virus preservation solution in the disposable virus sampling tube. 400
(2) Nucleic acid extraction: 60
(3) Using TaKaRa kits (RR391A and RR064A), we detected adenovirus (HADV), human respiratory syncytial virus (HRV), human rhinovirus (HRV), influenza A virus (FluA), influenza B virus (FluB) and human reference genes in the samples by real-time fluorescence quantitative PCR [16, 17, 18, 19]. The negative control was RNase Free ddH2O, and the primer probe was synthesized by Bio Engineering (Shanghai) Co., Ltd. The specific sequence and reaction conditions are shown in Table S1. The PCR detection conditions of DNA virus are as follows: [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 1; 95∘C for 30 s, [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 2; 95∘C for 5 s, [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 3; 95∘C for 30 s; Steps [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 2; and [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 3; were repeated for 45 cycles, and the temperature of step [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 3; was adjusted according to the annealing temperature in Table S1, and the reaction system is shown in Table S2. The detection conditions of RNA virus were as follows: [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 1; 42∘C for 5 min, [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 2; 95∘C for 5 s, [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 3; 95∘C for 10 s, [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 4; 60∘C for 30 s. Steps [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 2;, [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 3; and [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 4; were repeated for 45 cycles, the temperature of step [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 4; was adjusted according to the annealing temperature in Table S1, and the reaction system is shown in Table S3.
(4) The RNA nucleic acid extracted in step 2 was reverse transcribed into 30
(5) The highest positive detection rate of adenovirus was found after all samples were tested. The positive nucleic acid samples saved in step 4 were amplified by selecting some genes of adenovirus Hexon and using 2
(6) Nucleic acid electrophoresis: [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 1; Preparation of electrophoresis solution: 242 g of Tris and 37.2 g of Na2EDTA 2H2O were weighed by an electronic balance and dissolved in 800 ml RNase Free ddH2O, and then 57.1 ml of glacial acetic acid was added and stirred with a glass rod to make them fully dissolved. Finally, the volume was adjusted to 1 L with RNase Free ddH2O and stored at room temperature for later use. [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 2; Preparation of electrophoresis gel: 2 g of agarose was weighed and heated and dissolved in 100 ml of 0.5
2.2.3Adenovirus typing and construction of phylogenetic tree
The sequenced and successfully spliced nucleotide sequences were Blast analyzed on NCBI, and the sequences with high homology were selected as reference strains. Clustal W comparison was made for all sequences with MEGA7.0 (all parameters were selected by default values), and the phylogenetic tree was constructed by Neighbor-Joining (NJ) (Bootstrap Replications:1000 was set as the parameter, and the other parameters were all by default).
2.2.4Cell culture and adenovirus isolation
After the frozen cells were revived, the better-grown cells were selected for culture. After 6 h of culture, trypsin was added and the cells were digested in a 5% CO2 cell incubator at 37∘C for 2–3 min to prepare cell suspension. Then, the prepared cell suspension was absorbed and added to a 12-well plate with 2 ml per well, then numbered and transferred to 37∘C and cultured in a 5% CO2 cell incubator for 8–10 h. Adenovirus samples with ct value less than 30 were randomly selected and inoculated in odd-numbered wells, and serum-free DMEM medium was added to the other wells as the control group. After inoculation with virus, the 12-well plate was placed in a 5% CO2 cell incubator at 37∘C for 96 h, during which the cells were removed every 12 h and observed under a microscope to observe whether they were diseased or not. If the cells were found to be diseased, they were immediately placed in a freezing tube for temporary storage in a refrigerator at
2.2.5Extraction of total nucleic acids
Nucleic acids in the virus solution obtained in 1.2.4 were extracted by the Kunpeng ArchiPure12 Automated Nucleic Acid Extractor. 200 ml of viral liquid was aspirated in ArchiPure Universal Magnetic Bead-Based Rapid Viral DNA/RNA Extraction Kit (RGP-261002-48), 20
2.2.6Full-length sequencing of penton, hexon and fibrin genes
It can be seen from the adenovirus typing results that three strains of adenovirus C1 were successfully isolated in this experiment. According to the literature query, three general primers of major structural genes of adenopathy virus C, namely, penton, hexon and fibrin, were selected and amplified by TaKaRa Prime STAR® Max DNA Polymera (RR 045a) kit [20]. The primer was synthesized by Beijing Tianyi Huiyuan Biotechnology Co., Ltd., and the primer sequence is shown in Table S7. The amplification conditions were as follows: [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 1; denaturation at 94∘C for 30 s, [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 2; annealing at 55∘C for 5 s, [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 3; elongation at 72∘C for 1 min, [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 4; final elongation at 72∘C for 5 min. Steps [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 1;, [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 2; and [baseline=(char.base)] [shape=circle,draw,inner sep=0.2pt] (char) 3; were repeated for 30 cycles, and the reaction system is shown in Table S8.
The amplified PCR products were sent to Beijing Tianyi Huiyuan Biotechnology Co., Ltd. for sequencing, and the sequenced sequences were spliced in two directions by using the DNAMAN software. Each sequenced sequence was ensured to have about 150 bp fragments superimposed during splicing. The penton, hexon and fibrin gene sequences of the reference strain were downloaded from NCBI. The three conserved domains were compared in CluastlW and analyzed for homology using BioEdit. Then, the three conserved domains of penton, hexon and friber were analyzed for amino acid variation using MegAlign with the original strain AF534906 of adenovirus C1 as the reference strain, and the phylogenetic tree was constructed with MEAG7.0.
2.3Data collection
Apart from the information related to laboratory testing of the samples, general clinical data and clinical laboratory examination data related to the patients were also collected. General clinical data include: gender, age, cough and sputum, modified British Medical Research Council (MMRC) score, fever, dizziness, abnormal symptoms of digestive tract, chest tightness and discomfort, palpitation (abnormal heart rate), respiratory rate, and inhaled glucocorticosteroid use. Laboratory examination data include: white blood cell count, neutrophil count, lymphocyte count, monocyte count, eosinophil count, basophil count, liver function and renal function.
2.4Statistical analysis
All included data in this study were statistically processed using SPSS 26.0 statistical software. The K-S method was employed for normality test. Measurement data meeting normality were expressed by (
3.Results
3.1Comparison of general clinical data between the two groups
The results showed that there were 532 patients in the infected group, including 279 males and 253 females, with an average age of 71.23
Table 1
Comparison of general clinical data
| Item | Infection group ( | Non-infection group ( | ||
| Sex (M/F) | 279/253 | 552/439 | 1.482 | 0.223 |
| Age (yrs, | 71.23 | 71.34 | 1.739 | 0.082 |
| Cough and sputum | 3.010 | 0.082 | ||
| Cough and sputum | 425 | 753 | ||
| Dry cough | 107 | 238 | ||
| MMRC grade | 0.039 | 0.844 | ||
| Grade 3 | 52 | 100 | ||
| Grade 4 | 480 | 891 | ||
| Fever (T | 266 | 290 | 64.219 | |
| Dizziness ( | 329 | 590 | 0.769 | 0.380 |
| Gastrointestinal anomalies ( | 334 | 584 | 2.145 | 0.143 |
| Chest tightness ( | 424 | 822 | 2.453 | 0.117 |
| Palpitations ( | 263 | 80 | 339.424 | |
| Respiratory rate ( | 2.770 | 0.250 | ||
| | 253 | 504 | ||
| 21–25 bpm | 173 | 282 | ||
| 16–20 bpm | 106 | 205 | ||
| Use of inhaled corticosteroids ( | 484 | 901 | 0.001 | 0.969 |
| White blood cells ( | 10.31 | 11.45 | 0.315 | 0.752 |
| Neutrophils ( | 6.60 | 7.19 | 0.586 | |
| Lymphocytes ( | 1.55 | 1.42 | 1.395 | 0.162 |
| Eosinophils ( | 0.42 | 0.41 | 0.191 | 0.847 |
| Monocytes ( | 1.03 | 1.27 | 0.587 | 0.557 |
| Basophils ( | 0.06 | 0.08 | 0.799 | |
| Liver dysfunction ( | 175 | 356 | 1.398 | 0.237 |
| Kidney dysfunction ( | 79 | 185 | 3.522 | 0.061 |
| Dyslipidemia ( | 98 | 396 | 73.279 | |
| Heart failure ( | 212 | 296 | 15.513 | |
| Blood gas analysis ( | 0.183 | 0.912 | ||
| Type 2 respiratory failure | 478 | 890 | ||
| Type 1 respiratory failure | 28 | 56 | ||
| Hypoxemia | 26 | 45 | ||
| Disease course (d, | 17.48 | 12.45 | 5.682 | |
| Prognosis ( | ||||
| Deterioration | 87 | 85 | ||
| Poor | 283 | 85 | ||
| Remission | 162 | 821 |
3.2Clinical laboratory examination data of the two groups
The results showed that the proportion of lipid metabolism abnormalities (18.42% VS 39.96%,
3.3Respiratory virus infections in those with AECOPD
Among the 532 patients in the infected group in this study, 59 were rhinovirus, accounting for 11.09%, 83 were human respiratory syncytial virus, accounting for 15.60%, 404 were adenovirus, accounting for 75.94%, 24 were H1N1 virus, accounting for 4.51%, and 25 were influenza B, accounting for 4.70%. Eleven cases were infected with three viruses, accounting for 2.07%, and 37 cases were infected with two viruses, accounting for 6.95%.
Figure 1.
Agarose gel electrophoresis of PCR amplified products of 31 samples of adenovirus hexon genes.
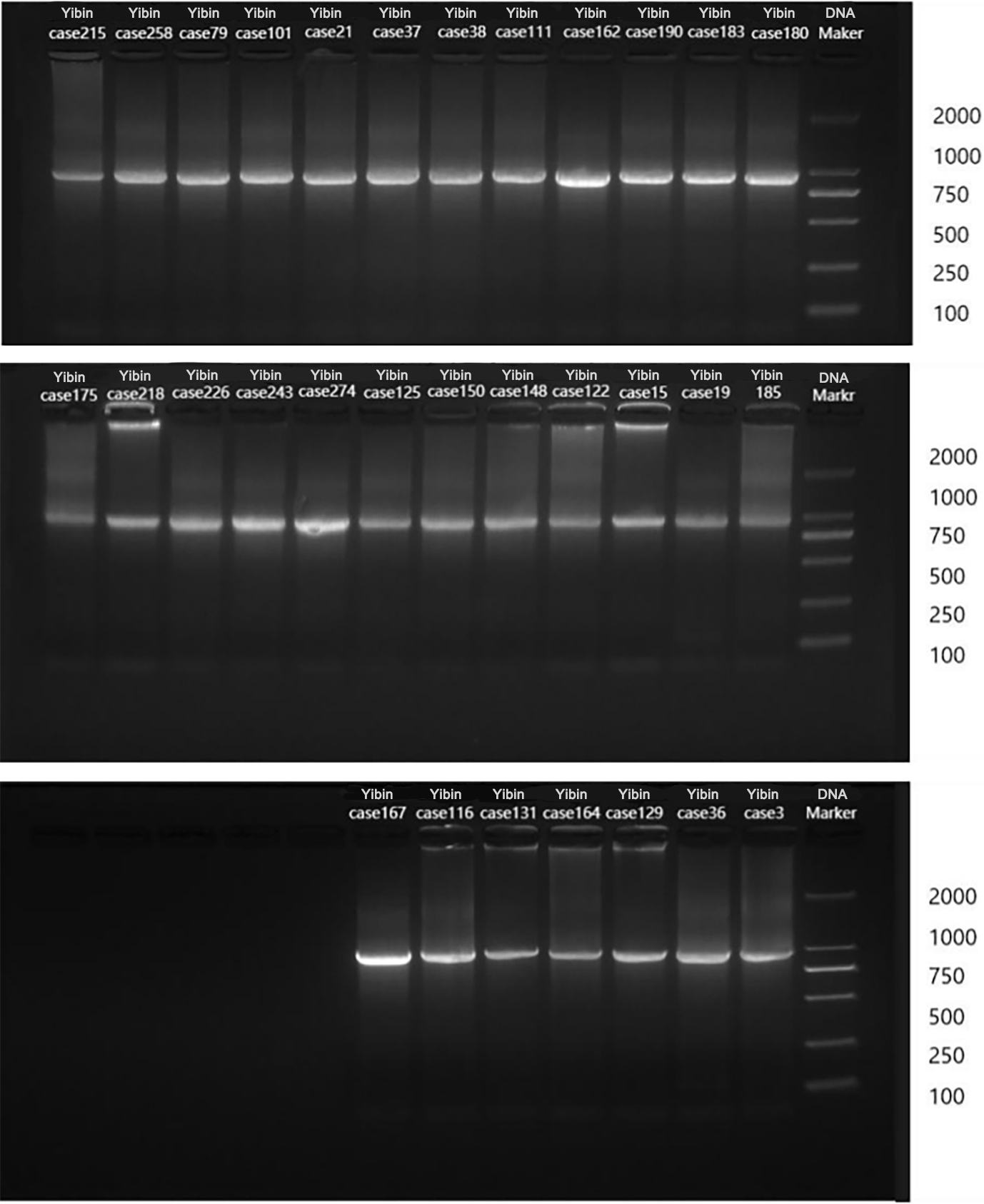
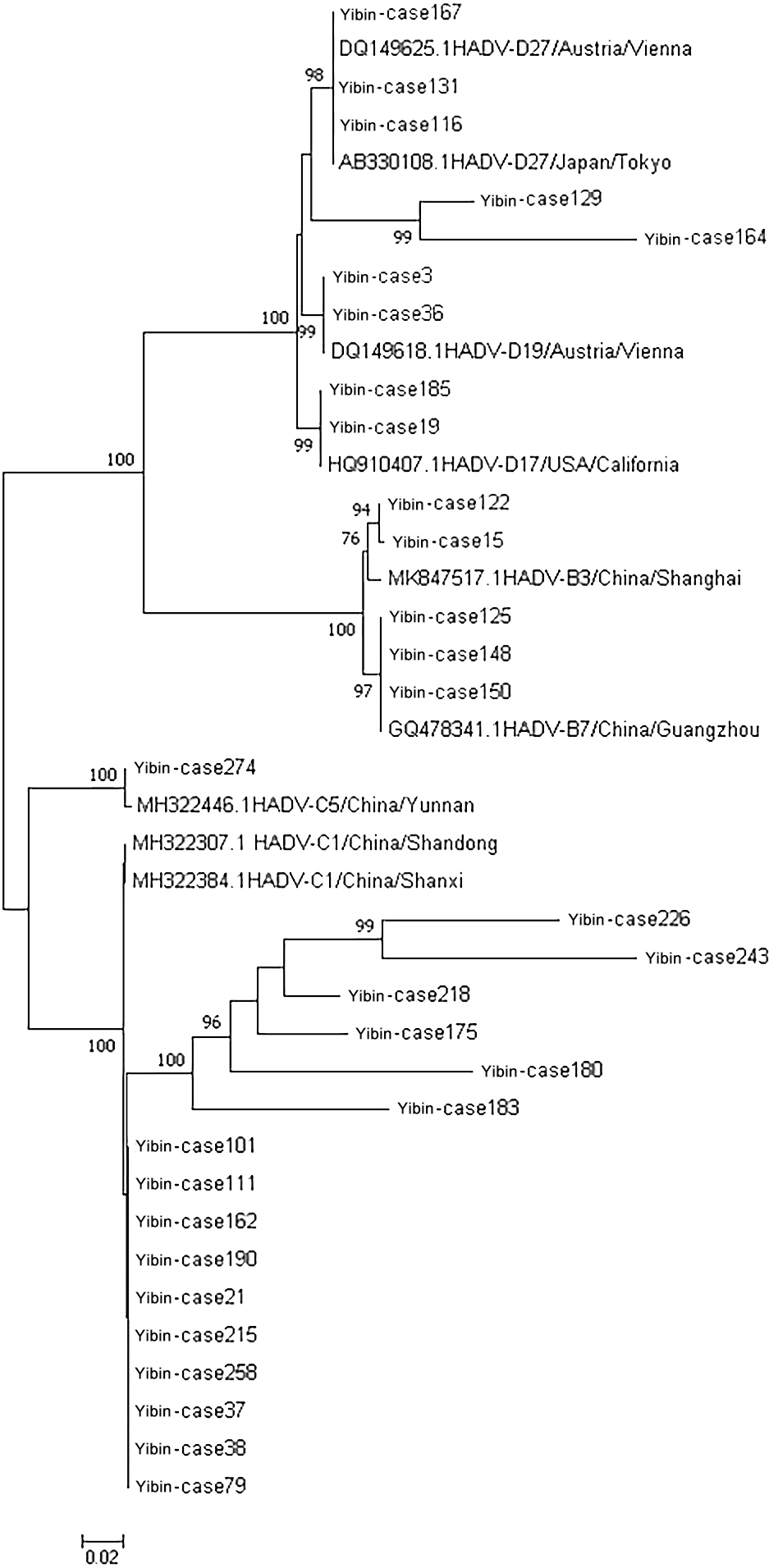
Figure 2.
Phylogenetic tree of 31 samples of adenovirus hexon genes.
3.4Adenovirus typing
Five common respiratory viruses were detected in throat swabs of 1523 patients with AECOPD, and 404 of them were positive for adenovirus. Subsequently, some fragments of hexon protein were amplified from adenovirus positive samples, and gel electrophoresis showed that 31 nucleic acids were amplified to a band of about 956 bp (see Fig. 1). The remaining PCR amplification products were sent to Beijing Tianyi Huiyuan Biotechnology Co., Ltd. for sequencing by Sanger’s method. The sequenced sequences were submitted to the National Center for Biotechnology Information (NCBI) for nucleotide sequence comparison. The results showed 16 ADV-C1 and 1 ADV-C5 for ADV-C type; 2 ADV-B3 and 3 ADV-B7 for ADV-B type; and 2 ADV-D17, 2 ADV-D19 and 5 ADV-D27 for ADV-D type. Among the 16 hexon sequences of ADV-C1 type, 10 sequences (named Yibin-case 258, 215, 79, 101, 21, 37, 38, 111, 162 and 190, respectively) were highly homologous to the sequences MH322384.1 reported in Shanxi, China, in 2007 and MH322307.1 reported in Shandong, China, in 2016, with a nucleotide sequence identity ranging from 99.24% to 99.79%. The remaining six ADV-C1 sequences (named Yibin-case 175, 180, 183, 218, 226 and 143, respectively) showed nucleotide sequence identity of 84.01%–98.10% with the reported sequences MH322384.1 reported in Shanxi, China, in 2007 MH322307.1 reported in Shandong, China, in 2016. An ADV-C5 sequence (named Yibin-case274) was highly homologous to the sequence MH322446.1 reported in Yunnan, China in 2014, with a nucleotide sequence identity of 99.48%. Two ADV-B3 sequences (named Yibin-case 15 and 122, respectively) were highly homologous to the sequence MK847517.1 reported in Shanghai, China in 2009, with a nucleotide sequence identity of 97.77% and 98.73%, respectively. Three ADV-B7 sequences (named Yibin-case 125, 148 and 150, respectively) were highly homologous to the sequence GQ478341.1 reported in Guangzhou, China in 2009, with a nucleotide sequence identity of 99.37%–99.68%. Nine ADV-D sequences were highly homologous to foreign reported sequences, among which two ADV-D17 sequences (named Yibin-case 19 and 185, respectively) were highly homologous to the sequence HQ910407.1 reported in California, USA, in 2011, with a nucleotide sequence identity of 99.47% and 99.58%, respectively. Two ADV-D19 sequences (named Yibin-case 3 and 36, respectively) were highly homologous to the sequence DQ149618.1 reported in Vienna, Austria in 2004, with a nucleotide sequence identity of 99.36% and 99.68%, respectively. Three of the five ADV-D27 sequences (named Yibin-case 116, 131 and 167, respectively) were highly homologous to sequences DQ149625.1 reported in Vienna, Austria in 2004 and AB330108.1 reported in Tokyo, Japan in 2007, with a nucleotide sequence identity of 99.79%, 99.68% and 99.23%, respectively; The other two sequences (named Yibin-case 129 and 164, respectively) were highly homologous to sequences DQ149625.1 reported in Vienna, Austria in 2004 and AB330108.1 reported in Tokyo, Japan in 2007, with a nucleotide sequence identity of 96.66% (see Fig. 2 for phylogenetic tree classification).
3.5Adenovirus typing
Throat swab samples with partial adenovirus hexon gene amplified and Ct value of real-time fluorescence quantitative PCR below 30 were selected to be inoculated on Hep-2 cells, respectively. After five generations of blind transmission, three strains of viruses were found to be pathological changes (see Fig. 3 for normal control and pathological changes). Subsequently, real-time fluorescence quantitative PCR of five respiratory viruses was performed again, and only adenovirus was positive after the assay; in addition, all adenovirus types were found to be adenovirus C1.
3.6Gene sequencing results of main structures of adenovirus (penton, hexon, friber)
Table 2
Adenovirus nucleotide homology and amino acid homology
| Penton | Nucleotides | Amino acids | ||||||
|---|---|---|---|---|---|---|---|---|
| 4 | 3 | 2 | 1 | 4 | 3 | 2 | 1 | |
| 4 | – | 0.996 | 0.995 | 0.997 | – | 0.993 | 0.993 | 0.996 |
| 3 | 0.996 | – | 0.994 | 0.996 | 0.993 | – | 0.990 | 0.994 |
| 2 | 0.995 | 0.994 | – | 0.996 | 0.993 | 0.990 | – | 0.995 |
| 1 | 0.997 | 0.996 | 0.996 | – | 0.996 | 0.994 | 0.995 | – |
| Hexo | Nucleotides | Amino acids | ||||||
| 4 | 3 | 2 | 1 | 4 | 3 | 2 | 1 | |
| 4 | – | 0.996 | 0.996 | 0.997 | – | 0.993 | 0.994 | 0.997 |
| 3 | 0.996 | – | 0.996 | 0.997 | 0.993 | – | 0.992 | 0.997 |
| 2 | 0.996 | 0.996 | – | 0.997 | 0.994 | 0.992 | – | 0.996 |
| 1 | 0.997 | 0.997 | 0.997 | – | 0.997 | 0.997 | 0.996 | – |
Note: 4 – adenovirus strain Yibin243; 3 – adenovirus strain Yibin162; 2 – adenovirus strain Yibin21; 1 – original adenovirus strain AF534906.1.
Figure 3.
Isolation of adenoviruses in Hep-2 cells vs. negative control. A: Negative control (10
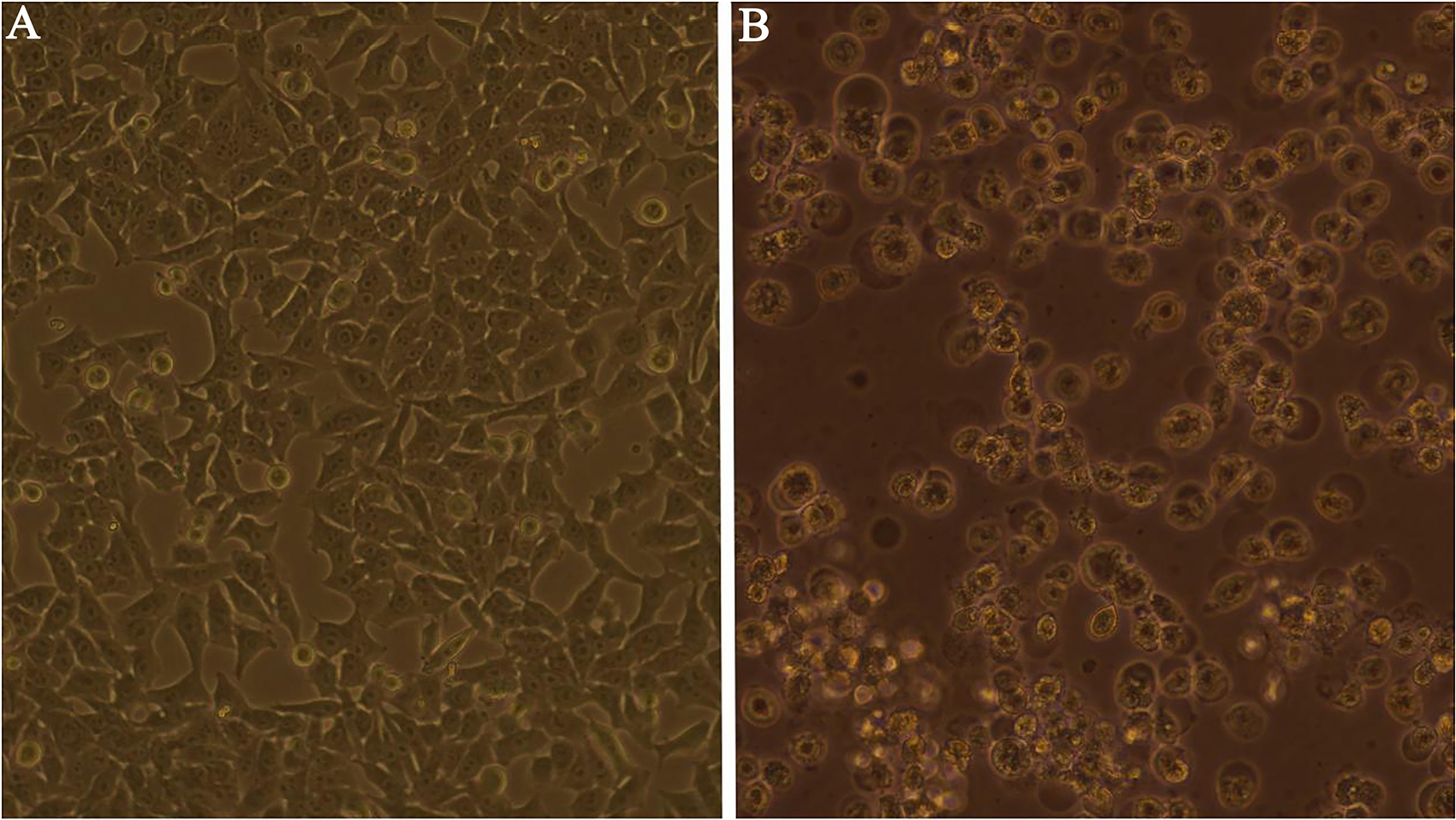
Figure 4.
Phylogenetic tree of adenovirus penton genes.
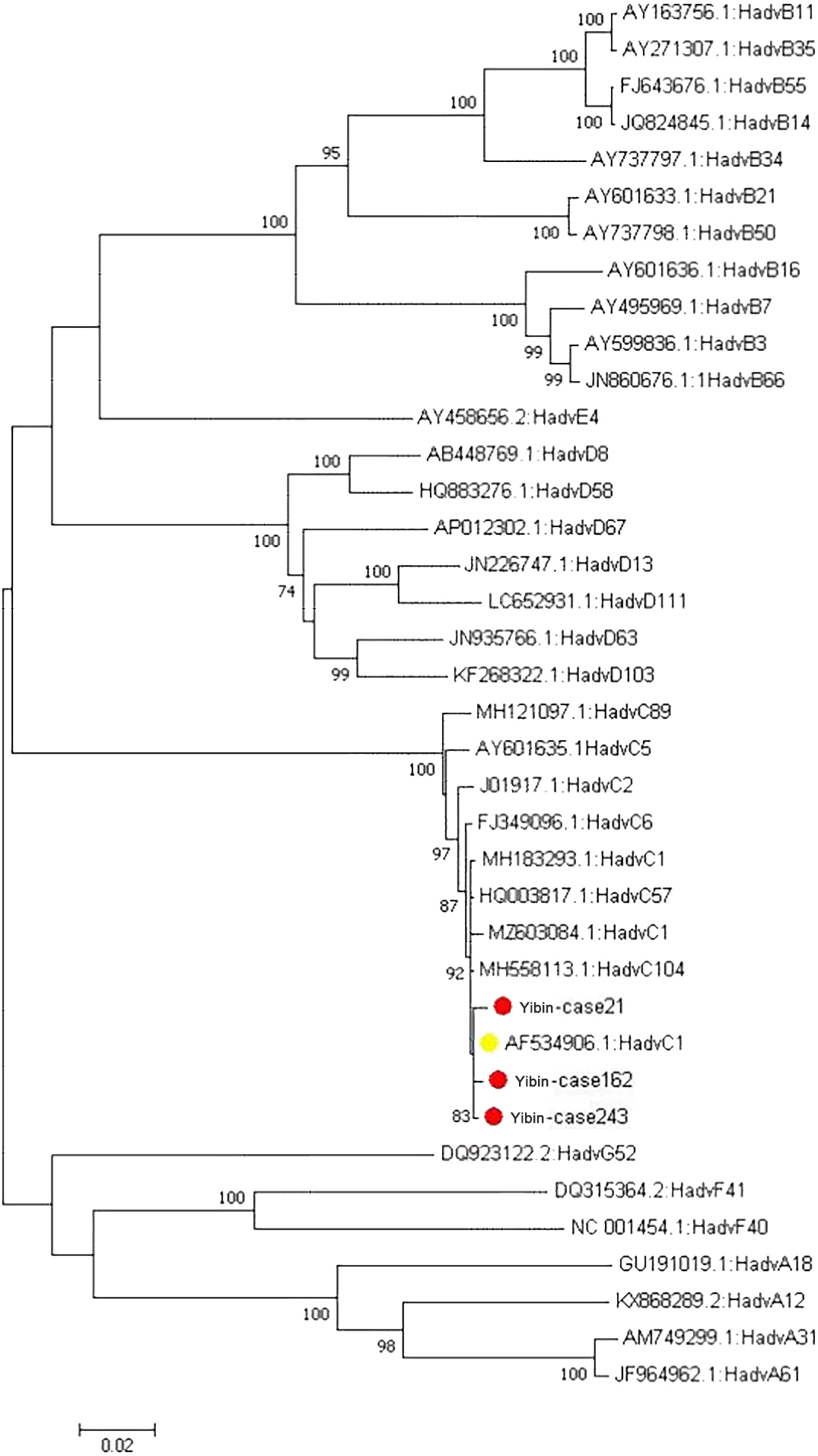
Table 3
Adenovirus gene mutation and amino acid variation
| No | Basic site | Base mutation | Amino acid site | Amino acid mutation |
|---|---|---|---|---|
| Pentongene mutation | ||||
| Yibin162 | 271 | T-G | 91 | F-A |
| Yibin162 | 272 | T-C | ||
| Yibin21 | 528 | C-A | Silent mutation | |
| Yibin243 | 536 | T-A | 179 | M-K |
| Yibin162 | 537 | G-C | 179 | M-I |
| Yibin162 | 544 | G-C | 182 | D-H |
| Yibin243 | 544 | G-A | 182 | D-N |
| Yibin162 | 919 | G-C | 307 | A-Q |
| Yibin162 | 920 | C-A | ||
| Yibin21 | 1261 | G-C | 421 | G-R |
| Yibin21 | 1349 | G-A | 450 | D-H |
| Yibin21 | 1389 | T-C | Silent mutation | |
| Yibin21 | 1430 | G-C | 477 | S-T |
| Yibin21 | 1665 | C-A | Silent mutation | |
| Hexon gene mutatio | ||||
| Yibin243 | 307 | A-T | 103 | I-F |
| Yibin21 | 890 | T-A | 297 | V-D |
| Yibin162 | 972 | G-A | Silent mutation | |
| Yibin162 | 1721 | T-A | 474 | V-P |
| Yibin162 | 1737 | A-G | 479 | F-L |
| Yibin21 | 1866 | T-C | Silent mutation | |
| Yibin243 | 1973 | A-G | 658 | Y-C |
| Yibin21 | 2845 | G-C | 949 | E-Q |
| Yibin243 | 2851 | G-C | 951 | V-L |
3.6.1Sequencing results of penton gene of adenovirus
The sequencing results were spliced in DNAMAN with the major structural genes of penton, hexon and friber. The sample numbers were named Yibin243, Yibin162 and Yibin21, respectively. Firstly, the three major structural gene sequences of three adenovirus strains were found to be highly homologous to the original strain of adenovirus C1 (AF534906) by Blast analysis on NCBI. Among them, the sequence identity of penton gene was 99.7%, 99.6% and 99.6%, respectively, and the amino acid similarity was 99.6%, 99.4% and 99.5%, respectively (Table 2). After comparing the penton gene sequences of adenovirus Yibin21 strain and the original strain (AF534906) on MegAlign, it was found that the mutations at three base sites, site 528, site 920 and site 1665, did not result amino acid mutation (synonymous mutation), and that site 1261 (G-C) resulted in the replacement of glycine at site 421 with arginine (G-R), site 1349 (G-A) resulted in the replacement of aspartic acid at site 450 with histidine (D-H), and site 1665 (G-C) resulted in the replacement of serine at site 477 with threonine. After comparing the penton gene sequences of adenovirus Yibin162 and the original strain (AF534906), six base site mutations were found, among which sites 271 (T-G) and 272 (T-C) jointly led to the replacement of phenylalanine at position 103 with alanine (F-A), sites 919 (G-C) and 920 (C-A) together resulted in the replacement of alanine at site 307 with glutamine (A-Q), sites 537 and 544 both (G-C) resulted in the replacement of methionine at site 179 with isoleucine (M-I), and site 182 resulted in the replacement of aspartic acid with histidine (D-H). After comparing the penton gene sequence of adenovirus Yibin243 and the original strain (AF534906), it was found that the base mutation (T-A) at site 536 resulted in the replacement of the corresponding methionine at site 179 with lysine (M-K), and the base mutation (G-A) at site 544 resulted in the replacement of the aspartic acid at site 182 with asparagine (Table 3). The penton gene sequence of three successfully isolated adenovirus strains and the penton gene sequence of adenovirus reference strain were constructed into a phylogenetic tree, and all of them were found on the same branch as the penton sequence of the original strain (AF534906) (see Fig. 4).
Table 4
Adenovirus friber nucleotide homology and amino acid homology
| Nucleotides | Amino acids | |||||||
|---|---|---|---|---|---|---|---|---|
| 4 | 3 | 2 | 1 | 4 | 3 | 2 | 1 | |
| 4 | – | 0.995 | 0.997 | 0.998 | – | 0.995 | 0.994 | 1 |
| 3 | 0.995 | – | 0.996 | 0.999 | 0.995 | – | 0.995 | 0.996 |
| 2 | 0.997 | 0.996 | – | 0.999 | 0.994 | 0.995 | – | 0.996 |
| 1 | 0.998 | 0.999 | 0.999 | – | 1 | 0.996 | 0.996 | – |
Note: 4 – adenovirus strain Yibin243; 3 – adenovirus strain Yibin162; 2 – adenovirus strain Yibin21; 1 – original adenovirus strain AF534906.1.
3.6.2Sequencing results of hexon gene of adenovirus
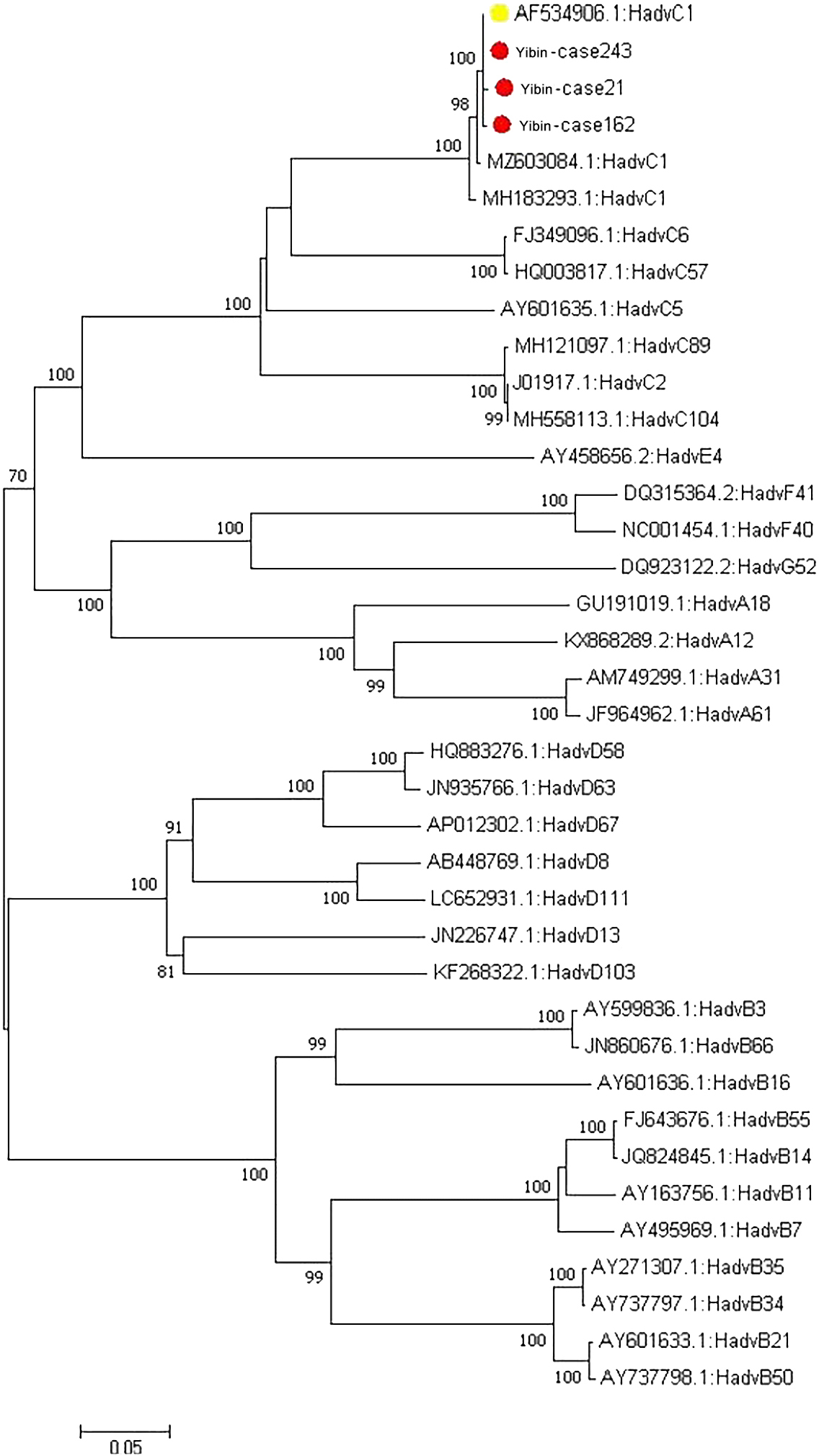
The nucleotide sequence of hexon gene of three adenovirus strains was 99.7% identical with that of the original strain, and the amino acid similarity was 99.7%, 99.7% and 99.6%, respectively (see Table 2). Comparison on MegAlign revealed that the base at site 1866 (T-C) in the Yibin21 strain and the base at site 972 (G-A) in the Yibin162 strain did not result in mutation of the corresponding encoded amino acid (synonymous mutation), that the base mutation at site 890 (T-A) in the Yibin21 strain resulted in the replacement of valine with aspartic acid at the corresponding site 297 (V-D), and that the base mutation at site 2845 (G-C) resulted in the replacement of glutamate with glutamine at the corresponding site (E-Q). The base mutation at site 1721 (T-A) in the hexon gene of the Yibin162 strain resulted in the replacement of valine with aspartic acid (V-P) at the corresponding site 474, and the base mutation at site 1737 (T-G) resulted in the replacement of phenylalanine with cysteamine (F-L) at the corresponding site 479. The base mutation (A-T) at site 307 of the hexon gene in the Yibin243 strain resulted in the replacement of the corresponding site 103 isoleucine with phenylalanine (I-F), the base mutation (G-C) at site 1973 resulted in the replacement of the corresponding tyrosine with cysteamine (Y-C), and the base mutation (G-C) at site 2851 resulted in the replacement of the corresponding valine with leucine (V-L) (Table 3). The penton gene sequence of the adenovirus and the penton gene sequence of adenovirus reference strain were constructed into a phylogenetic tree, and all of them were found on the same branch as the penton sequence of the original strain (AF534906) (see Fig. 5).
Figure 5.
Phylogenetic tree of adenovirus hexon genes.
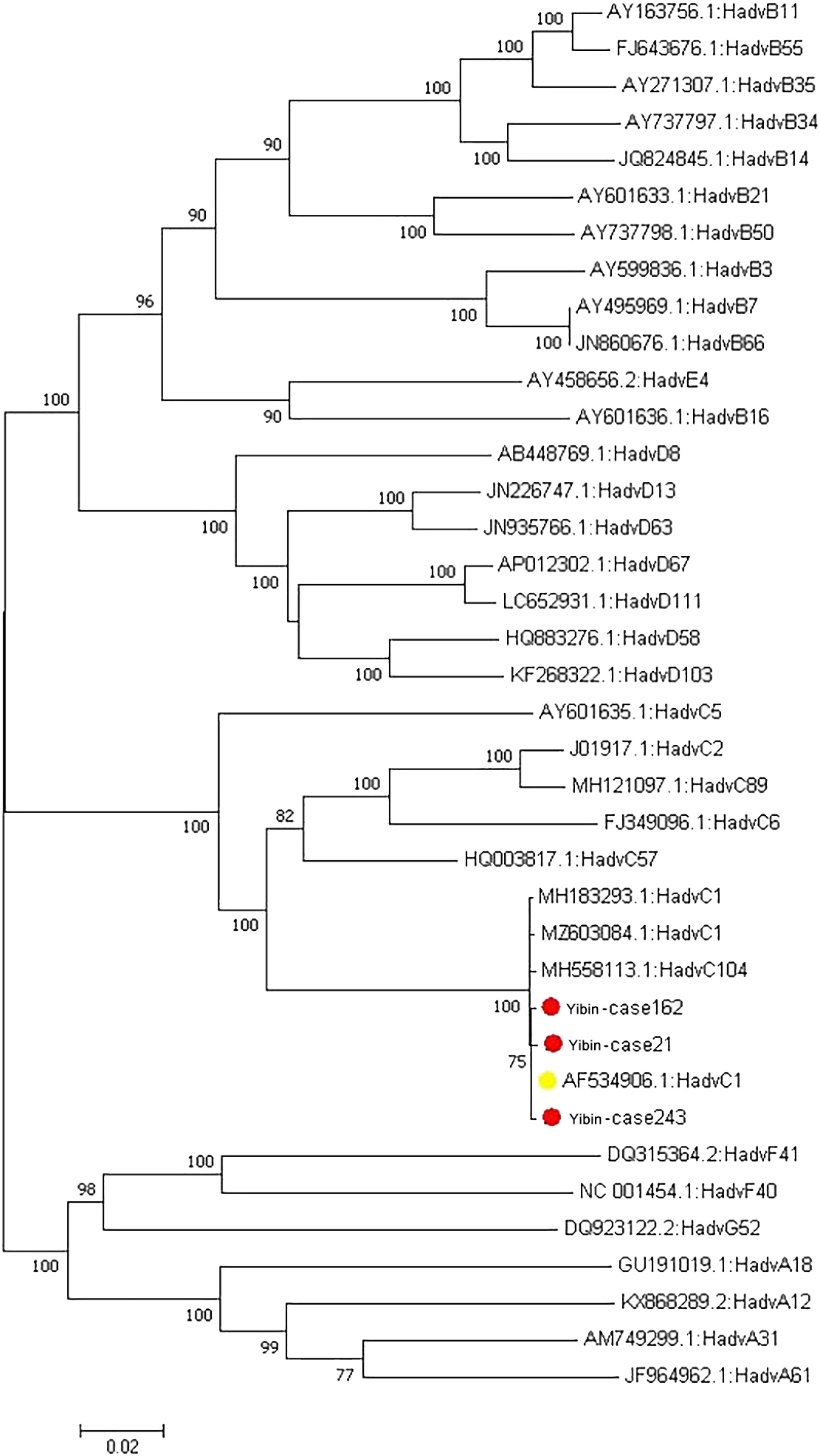
Figure 6.
Adenovirus friber gene sequencing results.
3.6.3Sequencing results of friber gene of adenovirus
The nucleotide sequences of the friber genes of the three adenovirus strains were 99.8%, 99.9% and 99.9% respectively, and the amino acids encoded by the friber genes of Yibin243 strain were 100% similar to those encoded by the original strain (AF534906.1), while those encoded by the friber genes of Yibin21 strain and Yibin162 strain were 100% similar to those encoded by the original strain (AF534906.1, Table 4). The base mutation (A-G) at site 1103 of Yibin21 strain resulted in the replacement of aspartic acid at position 368 with glycine (D-G), and the base mutations at sites 1412, 1585 and 1724 were all (C-A) resulted in the replacement of serine at site 471 with tyrosine (S-Y) and proline at site 529 with threonine (P), respectively. The base mutation at site 1226 (C-G) in the Yibin162 strain resulted in the replacement of the corresponding proline at site 409 with arginine (P-R), and the base mutation at site 1271 (T-G) resulted in the replacement of the corresponding valine at site 424 with glycine (V-G). No mutation (synonymous mutation) was seen in the amino group encoded by the base mutation at site 504 (T-C) in the Yibin243 strain (Table 5). The friber gene sequence of three successfully isolated adenovirus strains and the friber gene sequence of adenovirus reference strain were constructed into a phylogenetic tree, and all of them were found on the same branch as the friber sequence of the original strain (AF534906) (see Fig. 6).
Table 5
Adenovirus friber gene mutation and amino acid variation
| No | Basic site | Base mutation | Amino acid site | Amino acid mutation |
|---|---|---|---|---|
| Yibin243 | 504 | T-C | Silent mutation | |
| Yibin21 | 1103 | A-G | 368 | D-G |
| Yibin162 | 1226 | C-G | 409 | P-R |
| Yibin162 | 1271 | T-G | 424 | V-G |
| Yibin21 | 1412 | C-A | 471 | S-Y |
| Yibin21 | 1585 | C-A | 529 | P-T |
| Yibin21 | 1724 | C-A | 575 | T-N |
4.Discussion
Acute exacerbation of chronic obstructive pulmonary disease (AECOPD), the leading cause of hospitalization, can obviously affect the quality of life of patients, accelerate the progress of the disease, and significantly increase disability and mortality. Beeh [21], explored the correlation between acute exacerbation and hospitalization. During the whole study, it was known that 13.6% of the patients had recurrent attacks, of which 56.6% could not continue to control their symptoms through outpatient treatment, and the overall mortality increased by three times. The frequency of acute exacerbation would obviously affect the survival of patients. Meanwhile, the study found that patients with frequent exacerbations are more serious and the symptoms last longer, and they need to receive more pulmonary drug treatment [21], which adds to the economic burden of patients.
Patients with AECOPD have typical specific symptoms of COPD, such as cough and expectoration, asthma after exercise, and increased purulent sputum, as well as nonspecific symptoms such as self-consciousness, fatigue, fever, limb fatigue and discomfort, and insomnia. However, there are no clinically specific signs and biochemical markers available to distinguish the exacerbation of COPD caused by respiratory virus infection from that caused by non-viral infection [22]. Simon D Message et al. from Britain demonstrated in 2008 that rhinovirus infections mainly involved upper respiratory tract infection but rarely involved lower respiratory tract, and there was no specific change in general clinical examination [23]. In 2002, a comparative study of COPD inpatients by Rohde et al. in Germany pointed out that COPD inpatients with respiratory virus infection were more prone to fever than those without respiratory virus infection [24]. These results are similar to our findings, i.e., the fever of patients with AECOPD detected by virus infection is higher than that of those without virus infection, and the respiratory virus detection of patients with AECOPD with dyslipidemia is higher. The majority of patients with respiratory viral infections have palpitations, with a higher incidence of heart failure and a poorer prognosis than those without viral infections. In 2000, Greenberg et al. from the United States conducted a three-year longitudinal cohort study on COPD patients for the first time, which showed that respiratory virus infection would lead to an increase in the number, frequency and degree of exacerbation of COPD, and the hospitalization time and prognosis of patients were related to respiratory virus infection [25]. These results are similar to our findings, i.e., the course of disease of patients with respiratory virus infection (17.48
Adenovirus is the most common infection in the present study, which is different from other studies [26, 27]. The reasons for this are the seasonal time cycle and geographic variability of respiratory viruses, such as rhinovirus infection that causes common cold has been recorded all year round, influenza disease has increased explosively in winter compared with other seasons, and respiratory syncytial virus mainly broke out in the world from mid-October to mid-May of the following year [13, 28]. Influenza virus is the most common in Asia, while picornavirus is more common in Australia, Europe and North America [14].
In this study, 31 sequences were successfully sequenced and typed after amplification, which were similar to the serotypes reported at home and abroad that caused severe pneumonia [29, 30, 31]. Since 1975, Jim Williams et al. in the United States first proposed that adenovirus would continue to recombine and evolve, leading to the emergence of new serotypes [32], there have been many studies on adenovirus recombination. Last year, Joyce Odeke Akello et al. in Switzerland found that direct adenovirus, including penton protein, hexon protein and fibrin, were susceptible to amino acid variation and recombination on three major structural genes due to base insertions, substitutions, and deletions in the three major structural genes of direct adenoviruses [33]. This conclusion is consistent with the early inference of Japanese scholars that once the adenovirus has base mutation, amino acid mutation and genome recombination, the site of invasion on cell tissue will be widened, and the invasive ability will be enhanced, which will lead to a greater risk of life [34]. In our study, we retrospectively analyzed the clinical data of three patients with base and amino acid mutations and found that all of them were patients with type II respiratory failure and required ventilator maintenance therapy, with a very poor therapeutic prognosis. This is consistent with the conclusion put forward by Japanese scholars that the pathogenicity of adenovirus increases after base mutation and amino acid mutation.
Nevertheless, some limitations also exist in this study. First of all, this study is a retrospective analysis, which may be flawed in terms of the criteria for inclusion of subjects, selection of clinical data, and processing of data. Secondly, the success rate of virus isolation was much lower than the positive rate of PCR and later we will optimize the virus isolation and culture in the study step. The subjects included in this study are only limited to patients in our hospital, and the results may be biased. In response to this, more samples need to be included in subsequent studies to conduct in-depth research on acute exacerbation of COPD, and the data should be improved to provide a more accurate and realistic basis for more effective treatment.
5.Conclusion
To sum up, respiratory virus infection will lead to the deterioration of patients with AECOPD, accompanied by longer treatment cycle and poor prognosis. Patients with AECOPD who are also infected with respiratory viruses, such as adenovirus and respiratory syncytial virus, are more common. Screening for common respiratory viruses is necessary in the clinical treatment of AECOPD. This will be beneficial to early intervention of AECOPD complicated with respiratory virus, avoid further deterioration and provide effective data support for rehabilitation treatment of patients with COPD.
Conflict of interest
None to report.
Supplementary data
The supplementary files are available to download from http://dx.doi.org/10.3233/THC-240010.
References
[1] | Christenson SA, Smith BM, Bafadhel M, Putcha N. Chronic obstructive pulmonary disease. Lancet. (2022) Jun 11; 399: (10342): 2227-2242. doi: 10.1016/S0140-6736(22)00470-6. Epub 2022 May 6. PMID: 35533707. |
[2] | Hajian B, De Backer J, Vos W, van Geffen WH, De Winter P, Usmani O, Cahn T, Kerstjens HA, Pistolesi M, De Backer W. Changes in ventilation-perfusion during and after an COPD exacerbation: an assessment using fluid dynamic modeling. Int J Chron Obstruct Pulmon Dis. (2018) Mar 6; 13: : 833-842. doi: 10.2147/COPD.S153295. |
[3] | Vogelmeier CF, Román-Rodríguez M, Singh D, Han MK, Rodríguez-Roisin R, Ferguson GT. Goals of COPD treatment: Focus on symptoms and exacerbationsRespir. Med. (2020) May; 166: : 105938. doi: 10.1016/j.rmed.2020.105938. Epub 2020 Mar 21. |
[4] | Poon J, Campos M, Foronjy RF, et al. Cigarette smoke exposure reduces leukemia inhibitory factor levels during respiratory syncytial viral infection. (2019) ; 14: : 1305-1315. doi: 10.2147/COPD.S196658. |
[5] | Falsey AR, Walsh EE, Esser MT, et al. Respiratory syncytial virus–associated illness in adults with advanced chronic obstructive pulmonary disease and/or congestive heart failure. Journal of Medical Virology. (2019) ; 91: (1): 65-71. doi: 10.1002/jmv.25285. |
[6] | Gimenes JA, Srivastava V, ReddyVari H, et al. Rhinovirus-induces progression of lung disease in a mouse model of COPD via IL-33/ST2 signaling axis. Clinical Science. (2019) ; 133: (8): 983-996. doi: 10.1042/CS20181088. |
[7] | Saetta M, Di Stefano A, Turato G, et al. CD8+ T-lymphocytes in peripheral airways of smokers with chronic obstructive pulmonary disease. Am J Respir Crit Care Med. (1998) ; 157: (3 Pt 1): 822-6. doi: 10.1164/ajrccm.157.3.9709027. |
[8] | Hua HS, Wen HC, Weng CM, Lee HS, Chen BC, Lin CH. Histone deacetylase 7 mediates endothelin-1-induced connective tissue growth factor expression in human lung fibroblasts through p300 and activator protein-1 activation. J Biomed Sci. (2021) May 19; 28: (1): 38. doi: 10.1186/s12929-021-00735-5. |
[9] | Keicho N, Elliott WM, Hogg JC, et al. Adenovirus E1A gene dysregulates ICAM-1 expression in transformed pulmonary epithelial cells. Am J Respir Cell Mol Biol. (1997) ; 16: (1): 23-30. doi: 10.1165/ajrcmb.16.1.8998075. |
[10] | Moriyama M, Hugentobler WJ, Iwasaki A. Seasonality of Respiratory Viral Infections. Annu Rev Virol. (2020) Sep 29; 7: (1): 83-101. doi: 10.1146/annurev-virology-012420-022445. |
[11] | Nam HH, Ison MG. Respiratory syncytial virus infection in adults. BMJ. (2019) Sep 10; 366: : l5021. doi: 10.1136/bmj.l5021. PMID: 31506273. |
[12] | Abedi GR, Watson JT, Nix WA, et al. Enterovirus and Parechovirus Surveillance – United States, 2014-2016. MMWR Morb Mortal Wkly Rep. (2018) ; 67: (18): 515-518. doi: 10.15585/mmwr.mm6718a2. |
[13] | Li ZJ, Zhang HY, Ren LL, et al. Chinese Centers for Disease Control and Prevention (CDC) Etiology of Respiratory Infection Surveillance Study Team. Etiological and epidemiological features of acute respiratory infections in China. Nat Commun. (2021) Aug 18; 12: (1): 5026. doi: 10.1038/s41467-021-25120-6. |
[14] | Stolz D, Papakonstantinou E, Grize L, Schilter D, Strobel W, Louis R, Schindler C, Hirsch HH, Tamm M. Time-course of upper respiratory tract viral infection and COPD exacerbationEur. Respir J. (2019) Oct 10; 54: (4): 1900407. doi: 10.1183/13993003.00407-2019. Print 2019 Oct.PMID: 31391222. |
[15] | Vogelmeier CF, Criner GJ, Martinez FJ, et al. Global strategy for the diagnosis, management, and prevention of chronic obstructive lung disease 2017 Report. GOLD executive summary. Am J Respir Crit Care Med. (2017) ; 195: (5): 557-582. doi: 10.1164/rccm.201701-0218PP. |
[16] | Herbert ZT, Kershner JP, Butty VL, Thimmapuram J, Choudhari S, Alekseyev YO, Fan J, Podnar JW, Wilcox E, Gipson J, Gillaspy A, Jepsen K, BonDurant SS, Morris K, Berkeley M, LeClerc A, Simpson SD, Sommerville G, Grimmett L, Adams M, Levine SS. Cross-site comparison of ribosomal depletion kits for Illumina RNAseq library construction. BMC Genomics. (2018) Mar 15; 19: (1): 199. doi: 10.1186/s12864-018-4585-1. |
[17] | Heinicke F, Zhong X, Zucknick M, Breidenbach J, Sundaram AYM, Flåm TS, Leithaug M, Dalland M, Farmer A, Henderson JM, Hussong MA, Moll P, Nguyen L, McNulty A, Shaffer JM, Shore S, Yip HK, Vitkovska J, Rayner S, Lie BA, Gilfillan GD. Systematic assessment of commercially available low-input miRNA library preparation kits. RNA Biol. (2020) Jan; 17: (1): 75-86. doi: 10.1080/15476286.2019.1667741. |
[18] | Kim KJ, Yun SG, Cho Y, Nam MH, Lee CK. Usefulness of Combining Sputum and Nasopharyngeal Samples for Viral Detection by Reverse Transcriptase PCR in Adults Hospitalized with Acute Respiratory IllnessMicrobiol. Spectr. (2022) Dec 21; 10: (6): e0277522. doi: 10.1128/spectrum.02775-22. Epub 2022 Nov 14. |
[19] | Heim A, Ebnet C, Harste G, et al. Rapid and quantitative detection of human adenovirus DNA by real-time PCR. J Med Virol, (2003) ; 70: (2): 228-39. PMID: 12696109. |
[20] | Ishiko H, Shimada Y, Konno T, et al. Novel human adenovirus causing nosocomial epidemic keratoconjunctivitis. J Clin Microbiol, (2008) ; 46: (6): 2002--8. doi: 10.1128/JCM.01835-07. |
[21] | Beeh KM, Glaab T, Stowasser S, et al. Characterisation of exacerbation risk and exacerbator phenotypes in the POET-COPD trial. Respir Res. (2013) ; 14: : 116. doi: 10.1186/1465-9921-14-116. |
[22] | Tiwari A, Gautam AS, Pandey SK, Singh S, Singh RK. The role of RIPK1 in chronic obstructive pulmonary disease. Drug Discov Today. (2024) May 11: 104020. doi: 10.1016/j.drudis.2024.104020. |
[23] | Message SD, Laza-Stanca V, Mallia P, et al. Rhinovirus-induced lower respiratory illness is increased in asthma and related to virus load and Th1/2 cytokine and IL-10 production. Proc Natl Acad Sci USA. (2008) ; 105: (36): 13562-7. doi: 10.1073/pnas.0804181105. |
[24] | Rohde G, Wiethege A, Borg I, et al. Respiratory viruses in exacerbations of chronic obstructive pulmonary disease requiring hospitalisation: a case-control study. Thorax. (2003) ; 58: (1): 37-42. doi: 10.1136/thorax.58.1.37. |
[25] | Greenberg SB, Allen M, Wilson J, et al. Respiratory viral infections in adults with and without chronic obstructive pulmonary disease. Am J Respir Crit Care Med. (2000) ; 162: (1): 167-73. doi: 10.1164/ajrccm.162.1.9911019. |
[26] | Teper A, Colom AJ, Schubert R, Jerkic PS. Update in postinfectious bronchiolitis obliterans. Pediatr Pulmonol. (2023) Jun 28. doi: 10.1002/ppul.26570. |
[27] | Satia I, Cusack R, Greene JM, O’Byrne PM, Killian KJ, Johnston N. Prevalence and contribution of respiratory viruses in the community to rates of emergency department visits and hospitalizations with respiratory tract infections, chronic obstructive pulmonary disease and asthma. PLoS One. (2020) Feb 6; 15: (2): e0228544. doi: 10.1371/journal.pone.0228544. |
[28] | Yu J, Liu C, Xiao Y, et al. Respiratory Syncytial Virus Seasonality, Beijing, China, 2007-2015. Emerg Infect Dis. (2019) ; 25: (6): 1127-1135. doi: 10.3201/eid2506.180532. |
[29] | Heim A. [Adenovirus infections]. Monatsschr Kinderheilkd. (2020) ; 168: (6): 514-523. doi: 10.1007/s00112-020-00909-9. |
[30] | Pscheidt VM, Gregianini TS, Martins LG, et al. Epidemiology of human adenovirus associated with respiratory infection in southern Brazil. Rev Med Virol. (2021) ; 31: (4): e2189. doi: 10.1002/rmv.2189. |
[31] | Yao LH, Wang C, Wei TL, et al. Human adenovirus among hospitalized children with respiratory tract infections in Beijing, China, 2017–2018. Virol J. (2019) ; 16: (1): 78. doi: 10.1186/s12985-019-1185-x. |
[32] | Williams J, Grodzicker T, Sharp P, et al. Adenovirus recombination: physical mapping of crossover events. Cell. (1975) ; 4: (2): 113-9. doi: 10.1016/0092-8674(75)90117-8. |
[33] | Akello JO, Kamgang R, Barbani MT, et al. Genomic analyses of human adenoviruses unravel novel recombinant genotypes associated with severe infections in pediatric patients. Sci Rep. (2021) ; 11: (1): 24038. doi: 10.1038/s41598-021-03445-y. |
[34] | Kaneko H, Aoki K, Ohno S, et al. Complete genome analysis of a novel intertypic recombinant human adenovirus causing epidemic keratoconjunctivitis in Japan. J Clin Microbiol. (2011) ; 49: (2): 484-90. doi: 10.1128/JCM.01044-10. |


