
Glucocerebrosidase Activity is Reduced in Cryopreserved Parkinson’s Disease Patient Monocytes and Inversely Correlates with Motor Severity
Abstract
Background:
Reduced activity of lysosomal glucocerebrosidase is found in brain tissue from Parkinson’s disease patients. Glucocerebrosidase is also highly expressed in peripheral blood monocytes where its activity is decreased in Parkinson’s disease patients, even in the absence of GBA mutation.
Objective:
To measure glucocerebrosidase activity in cryopreserved peripheral blood monocytes from 30 Parkinson’s disease patients and 30 matched controls and identify any clinical correlation with disease severity.
Methods:
Flow cytometry was used to measure lysosomal glucocerebrosidase activity in total, classical, intermediate, and non-classical monocytes. All participants underwent neurological examination and motor severity was assessed by the Movement Disorders Society Unified Parkinson’s Disease Rating Scale.
Results:
Glucocerebrosidase activity was significantly reduced in the total and classical monocyte populations from the Parkinson’s disease patients compared to controls. GCase activity in classical monocytes was inversely correlated to motor symptom severity.
Conclusion:
Significant differences in monocyte glucocerebrosidase activity can be detected in Parkinson’s disease patients using cryopreserved mononuclear cells and monocyte GCase activity correlated with motor features of disease. Being able to use cryopreserved cells will facilitate the larger multi-site trials needed to validate monocyte GCase activity as a Parkinson’s disease biomarker.
INTRODUCTION
Pathogenic variations in GBA1, the gene encoding the lysosomal hydrolase glucocerebrosidase (GCase), increase the risk of developing neurodegenerative Parkinson’s disease (PD) [1, 2]. Exactly how GBA1 contributes to PD risk is still unclear; however, missense mutations that impair the enzymatic activity of the encoded enzyme GCase can result in lysosomal dysfunction and the accumulation of the pathological PD protein alpha-synuclein (for recent reviews see [3–5]). Importantly, reduced GCase activity has also been observed in association with alpha-synuclein pathology in brain tissue from patients even without GBA1 variations [6–9]. This suggests that impaired GCase activity may be a common mechanism in PD and consequently there is much interest in targeting the GCase enzyme as a potential therapeutic strategy for PD [10, 11].
In addition to the central nervous system, GCase is also highly expressed in the periphery, particularly in immune cells of myeloid lineage such as monocytes and macrophages. Large scale studies using whole blood samples have indicated that peripheral GCase activity is reduced in PD patients, including those without GBA1 variations, which may therefore offer utility for predicting future clinical outcomes and/or the stratification of patients into clinical trials [12–14]. However, heterogeneity in the cell populations comprising whole blood can complicate the measurement of GCase activity. To assess GCase activity in more homogenous immune cell populations, we recently employed a flow cytometry approach to measure GCase activity in the immune cell subsets that comprise peripheral blood mononuclear cells (PBMCs). This approach indicated a significant reduction in GCase activity in idiopathic PD patient monocytes, but not in the lymphocytes that comprise the majority of PBMCs [15]. These results suggest that single cell approaches may provide improved resolution for assessing GCase activity in patient populations.
However, one limitation of our recent study was that the flow cytometry assessment of GCase activity was performed using freshly isolated blood samples, collected on the day that participants presented to the clinic. While convenient for a small study, this approach would be unfeasible for larger, multi-center studies wishing to utilize monocyte GCase activity as a PD biomarker. To address this problem, in the current study, we have cryopreserved PBMCs from control and idiopathic PD patients. We aimed to determine whether monocyte GCase activity was still reduced in PD patient monocytes compared to age matched controls from cryopreserved specimens. In addition, we sought to correlate GCase activity with motor severity scores to determine a potential role for this approach in future clinical trials.
METHODS AND MATERIALS
Participants
A total of 30 PD patients and 30 healthy control subjects were recruited with informed consent in accordance with applicable guidelines and regulations that were approved by the University of Sydney Human Research Ethics Committee (2016/363). The diagnosis of PD followed the Movement Disorders Society (MDS) criteria for clinically established PD [16]. Exclusion criteria were a first degree relative with PD and the current use of non-steroidal anti-inflammatory medication. Control subjects were age and sex matched to PD patients, with 25 out of 30 controls having identified as being a spouse, sibling or partner, with a further 5 controls that had no connection to the participants. Recruitment occurred over a four-month period with both controls and PD patient samples being collected and processed on the same days by the same staff. Demographic data, which comprised age, sex, time since diagnosis, MDS Unified Parkinson Disease Rating Scale part 3 (MDS-UPDRS III) score, and the levodopa equivalent dose for PD patients receiving dopaminergic medication were recorded at the time of blood collection.
Isolation and cryopreservation of peripheral blood mononuclear cells from participants
Venous blood was collected into 8 mL sodium citrate Vacutainer Glass Cell Preparation Tubes (BD Biosciences), with the tubes kept upright following blood collection. Tubes were inverted 8 times prior to centrifugation to remix the sample, after which they were centrifuged at 1,800 x g for 20 min at room temperature. The PBMC layer was then transferred to a 15 ml conical tube, which was supplemented with room temperature phosphate buffered saline (PBS) to bring the volume to 15 ml. The PBMCs were centrifuged at 300 x g for 15 min at room temperature, after which the supernatant was aspirated and the PBMC pellet was resuspended in 10 ml of warmed (37°C) culture media (RPMI 1640 supplemented with 10% v/v heat inactivated, low endotoxin fetal bovine serum (FBS) and 1 X penicillin/streptomycin (all from Thermo Fisher Scientific). Cell count and viability were determined by using Trypan blue and an automated cell counter (Countess II-FL, Life Technologies). Cells were then pelleted via centrifugation at 300 x g for 10 min at room temperature, after which the supernatant was decanted and cells resuspended in pre-chilled (4°C) cryopreservation media (RPMI 1640 supplemented with 20% v/v FBS and 10% v/v dimethyl sulfoxide (DMSO) (Sigma-Aldrich). Cell suspensions were frozen at –80°C overnight in a Mr. Frosty Freezing Container (Thermo Fisher Scientific). Cryovials were then transferred into liquid nitrogen storage within one week and the storage time in liquid nitrogen was recorded in months.
Flow cytometry assessment of GCase activity
Cryopreserved PBMCs were retrieved from liquid nitrogen, thawed at 37°C, pelleted via centrifugation and resuspended in warmed (37°C) culture media. The maximum storage period of PBMCs in liquid nitrogen was four months. PBMCs were then incubated at 37°C in a 5% CO2 tissue culture incubator for 2 h. Following the 2 h recovery period, PBMCs from each subject were counted and 1×106 cells were transferred into each of two new 1.5 ml centrifuge tubes. One tube was treated with 1 mM of the GCase inhibitor Conduritol B epoxide (CBE) (Sigma-Aldrich), and the other treated with an equivalent volume of DMSO, for 1 h at 37°C. Following this, 0.75 mM of the lysosomal GCase substrate, 5-(Pentafluorobenzoylamino) Fluorescein Di-β-D-Glucopyranoside (PFB-FDGlu) (Thermo Fisher Scientific), was added to both tubes for 30 min at 37°C and GCase enzyme activity assessed, as we have published before [15]. To terminate the reaction, 1 ml of pre-chilled (4°C) fluorescence-activated cell sorting (FACS) buffer (1 X PBS, 1 mM EDTA, 25 mM HEPES, 1% heat inactivated FBS, pH 7.4) was added to tubes, and PBMCs were pelleted via centrifugation at 300 x g for 5 min at 4°C. To assess GCase activity in different monocyte subsets, PBMCs were resuspended in a master mix of FACS buffer containing PE-Cy7-conjugated anti-CD14, V450-conjugated anti-CD16, and PE-conjugated anti-HLA-DR monoclonal antibodies (BD Biosciences). Tubes were incubated for 30 min at 4°C to stain cell surface markers. PBMCs were then washed and resuspended in FACS buffer, after which they were passed through a cell strainer cap and kept on ice until data acquisition. Data were acquired on a BD FACSMelody instrument (BD Biosciences) and FCS files analysed using FlowJo software (Tree Star Inc.). Approximately 50,000 events were captured from each sample. Monocyte subsets were distinguished by their relative expression of cell surface markers; classical monocytes were CD14 + CD16-, intermediate monocytes were CD14 + CD16+, and nonclassical monocytes were CD14dim CD16 + . HLA-DR was incorporated into the gating strategy to identify and exclude NK cells and B cells from the analysis. Fluorescent signal resulting from PFB-FDGlu metabolism within each monocyte population was then quantified and expressed as an index, obtained by dividing the median fluorescence intensity (MFI) of cells treated without CBE by the MFI of cells treated with CBE.
GBA1 sequencing
Venous blood was collected from participants into 4 ml EDTA tubes (BD Biosciences) and genomic DNA was isolated using a QIAamp DNA Mini Kit (QIAGEN), according to manufacturers’ instructions. Sanger sequencing of GBA1 was performed using long-range PCR, as we have published before [6, 15].
Statistical analysis
Student’s t-test using Prism software (Graphpad) was used to compare between two groups with statistical significance defined as p < 0.05. General linear model multivariate analysis using SPSS (IBM) was also used to compare between groups whilst allowing for age, sex, percent of monocytes in PBMCs and storage time in months to be included as covariates. Statistical significance for multivariate analysis was accepted at p < 0.05 using the Wilks’ Lambda statistic. Partial correlation analyses were used to determine if there was a negative association between monocyte GCase activity and UPDRSIII using the same covariates as listed for the multivariate analysis. Spearman correlation was performed to determine any association between monocyte GCase activity and Dopamine equivalent daily dose. Graphs were created using Prism software version 7.0 (GraphPad) and show individual data points and the estimated marginal mean ± S.E.M.
RESULTS
Reduced GCase activity in cryopreserved PD patient monocytes
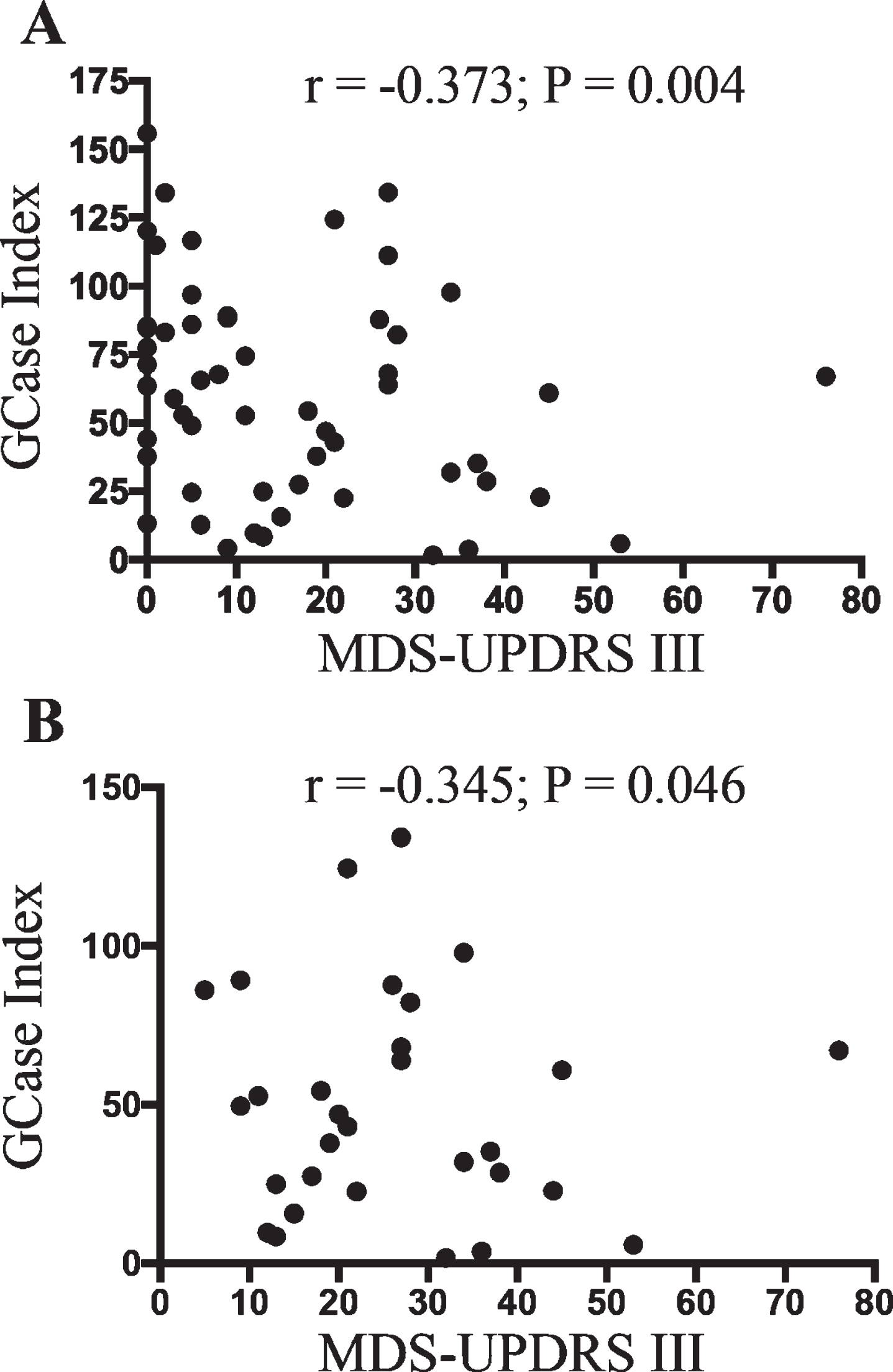
A total of 60 participants were recruited for the study. The control and PD groups were matched for age and sex, and as expected, PD patients had significantly higher scores on the MDS UPDRS III motor severity scale (Table 1). Sanger sequencing of the GBA1 gene revealed one PD patient with an N409S variant, who was excluded from further analysis. At the end of recruitment, cryopreserved PBMCs were used to assess GCase activity in total CD14 positive monocytes, as well as in different monocyte subsets using the expression of CD14 and CD16 to define classical, intermediate, and non-classical groups. The gating strategy is shown in Fig. 1. No significant differences were found between the control and PD groups for the percentage of total monocytes, or monocyte subpopulations in the blood samples (Table 1). Boxplot analysis did reveal that three control participants had percentages of classical and intermediate monocytes that were more than 3 times lower and higher respectively than the interquartile range. These three outliers were removed from further analysis. Consistent with previous work using freshly isolated cells, Student’s t-test revealed a significant reduction in the GCase activity index in cryopreserved total CD14 positive monocytes (p < 0.05, Fig. 2A). Using surface marker antibodies to split the total monocyte population into the different monocyte subsets revealed a significant decrease in the GCase activity index in classical monocytes (p < 0.05, Fig. 2B) from PD patients compared to controls, with no difference observed between the two groups in intermediate (Fig. 2C) and non-classical (Fig. 2D) monocytes. Multivariate analyses covarying for age, sex, the percent of monocytes in the PBMC sample and the storage time of the cryopreserved PBMCs was also performed. Neither age (p = 0.674), sex (p = 0.980) or the percent of monocytes in the blood (p = 0.059) had a significant effect on the GCase activity index, however storage time did (p = 0.022). After adjusting for the above covariates, the GCase activity index was still significantly reduced in PD patient total (p = 0.003, Fig. 2E) and classical (p = 0.004, Fig. 2F) monocytes. Employing raw MFI values from the metabolism of PFB-FDGlu for GBA activity, rather than the GBA index corrected for background signal, in the multivariate analysis with the same covariates resulted in a significant reduction in MFI in PD patients in total monocytes (GBA MFI; control = 9264±998, PD = 5545±979, p = 0.012), but not classical monocytes (GBA MFI; control = 9882±942, PD = 7405±924, p = 0.072). This highlights the utility of including a control for background signal such as the GCase inhibitor CBE. In this study CBE blocked 85±3% and 95±1% of the MFI signal from the metabolism of PFB-FDGlu in total and classical monocytes respectively, with no significant difference in the signal blocked between the PD and control groups. Interestingly, across the whole cohort, a one-way ANOVA revealed that GCase activity for intermediate monocytes was significantly lower than for the classical or non-classical subsets (p < 0.001, Fig. 3). To determine if reduced GCase activity is associated with worse motor symptoms assessed by UPDRSIII, partial correlation analysis was performed controlling for age, sex, percent of monocytes in the PBMCs and storage time. Across the whole cohort, a significant negative correlation between classical monocyte GCase activity and MDS-UPDRS III motor severity scores (r = –0.373; p = 0.004, Fig. 4A) but not for the intermediate (p = 0.363) or non-classical (p = 0.079) monocytes. When just the PD patients were included in the correlation analysis, a significant negative correlation between classical monocyte GCase activity and MDS-UPDRS III motor severity scores is still observed (r = –0.345; p = 0.046, Fig. 4B). Consistent with our previous study, there was no correlation between dopamine replacement medication and monocyte GCase activity (p > 0.8).
Table 1
Demographic data for PD patients and control subjects. Sixty participants in total were recruited for this study and the demographic data is shown. Data are presented as the mean±standard error of the mean. LEDD, L-Dopa daily equivalent dose; MDS-UPDRSIII, Movement Disorder Society Unified Parkinson’s Disease Rating scale part 3; PBMCs, peripheral blood mononuclear cells
| Parkinson’s disease | Control | |
| Number of participants | 30 | 30 |
| Age (y) | 68±1 | 68±1 |
| Male % | 73 | 66 |
| Time since diagnosis (y) | 6.7±0.80 | N/A |
| LEDD | 758±67 | N/A |
| MDS-UPDRS III | 26.3±15.1* | 3.9±5.5 |
| Total monocytes (% of PBMCs) | 26±9 | 21±10 |
| Classical monocytes (% total monocytes) | 65±21 | 70±16 |
| Intermediate monocytes (% total monocytes) | 22±18 | 18±15 |
| Non-classical monocytes (% total monocytes) | 9±5 | 9±6 |
Fig. 1
Flow cytometry gating strategy to distinguish classical, intermediate, and non-classical monocyte subsets and quantify GCase activity in participant PBMCs. PBMCs were treated with the lysosomal GCase substrate PFB-FDGlu after a preceding incubation with or without the GCase inhibitor CBE and stained with antibodies to identify monocyte subpopulations. Firstly, a singlet gate was created using FSC-A and FSC-H parameters (A), followed by a monocyte gate based on FSC-A and SSC-A parameters (B). CD14 positive cells were then selected (C), and fluorescent signal in the FL1-A channel, resulting from PFB-FDGlu metabolism by GCase, was assessed in this total monocyte population (D). Additionally, monocytes were further categorized into distinct subsets based on the expression of CD14 and CD16 (E), followed by a gate to exclude cells positive for CD16 but negative for HLA-DR, representing NK cells (F). Cells expressing high HLA-DR but low CD14, characteristic of B cells, were also excluded (G). Finally, monocyte subsets were distinguished on the basis of relative CD14 and CD16 expression levels (H), and GCase activity was quantified in classical (I), intermediate (J), and non-classical (K) monocytes. Raw GCase activity was expressed as an index, obtained from dividing the MFI of cells treated without CBE by the MFI of cells treated with CBE.
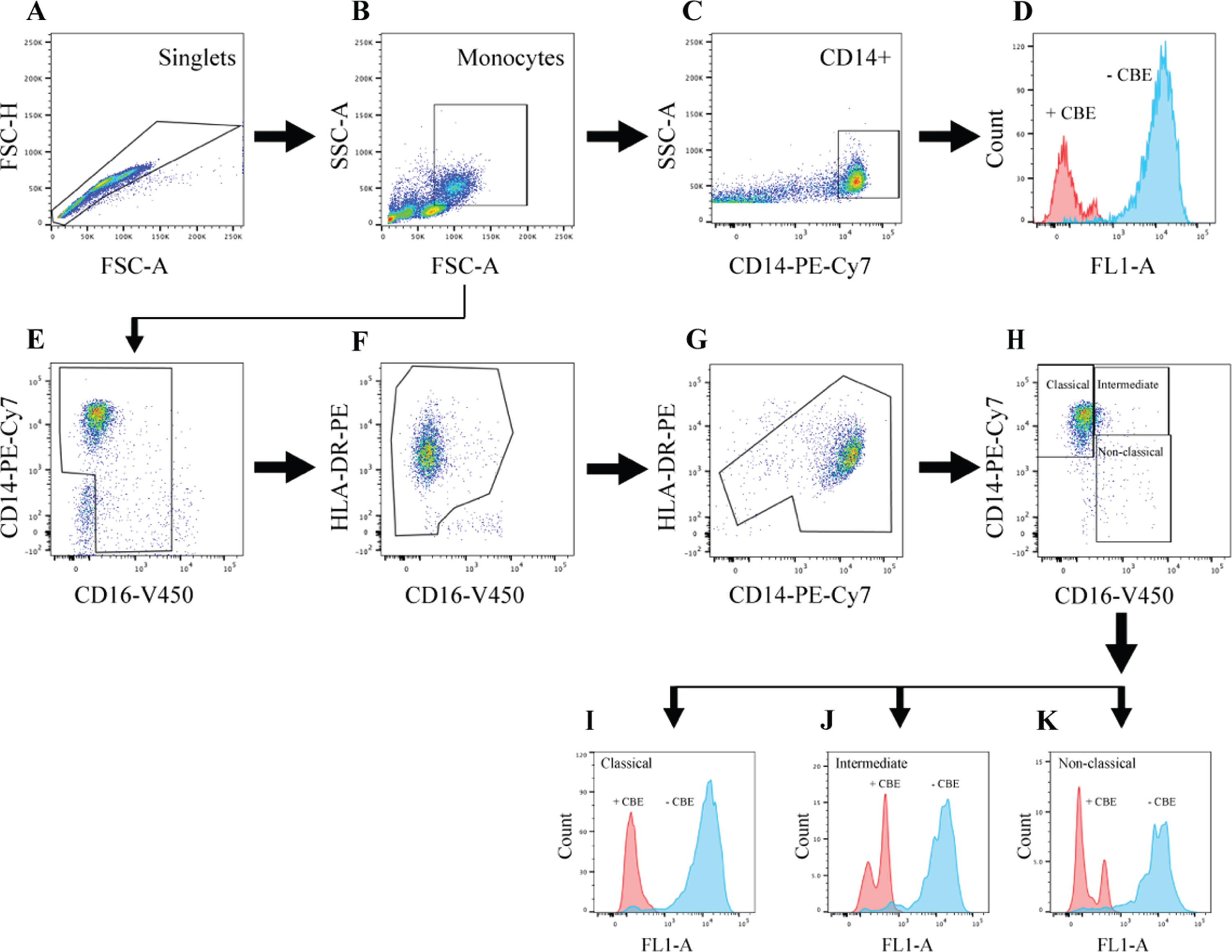
Fig. 2
Reduced GCase activity in PD patient monocytes. Student’s t-test was used to compare monocyte GCase activity between PD (n = 29) and control (n = 27) subjects in total (CD14+) (A), classical (CD14++CD16-) (B), intermediate (CD14 + CD16+) (C), and non-classical (CD14lowCD16+) (D) monocyte subsets. Data are presented as the mean±SEM with each dot representing an individual participant. Multivariate analysis covarying for age, sex, the percent of monocytes in peripheral blood mononuclear cells and mononuclear cell storage time in months showed a significant reduction in GCase activity in PD patient total (E) and classical (F) monocytes. Data are presented as the estimated marginal mean±SEM.
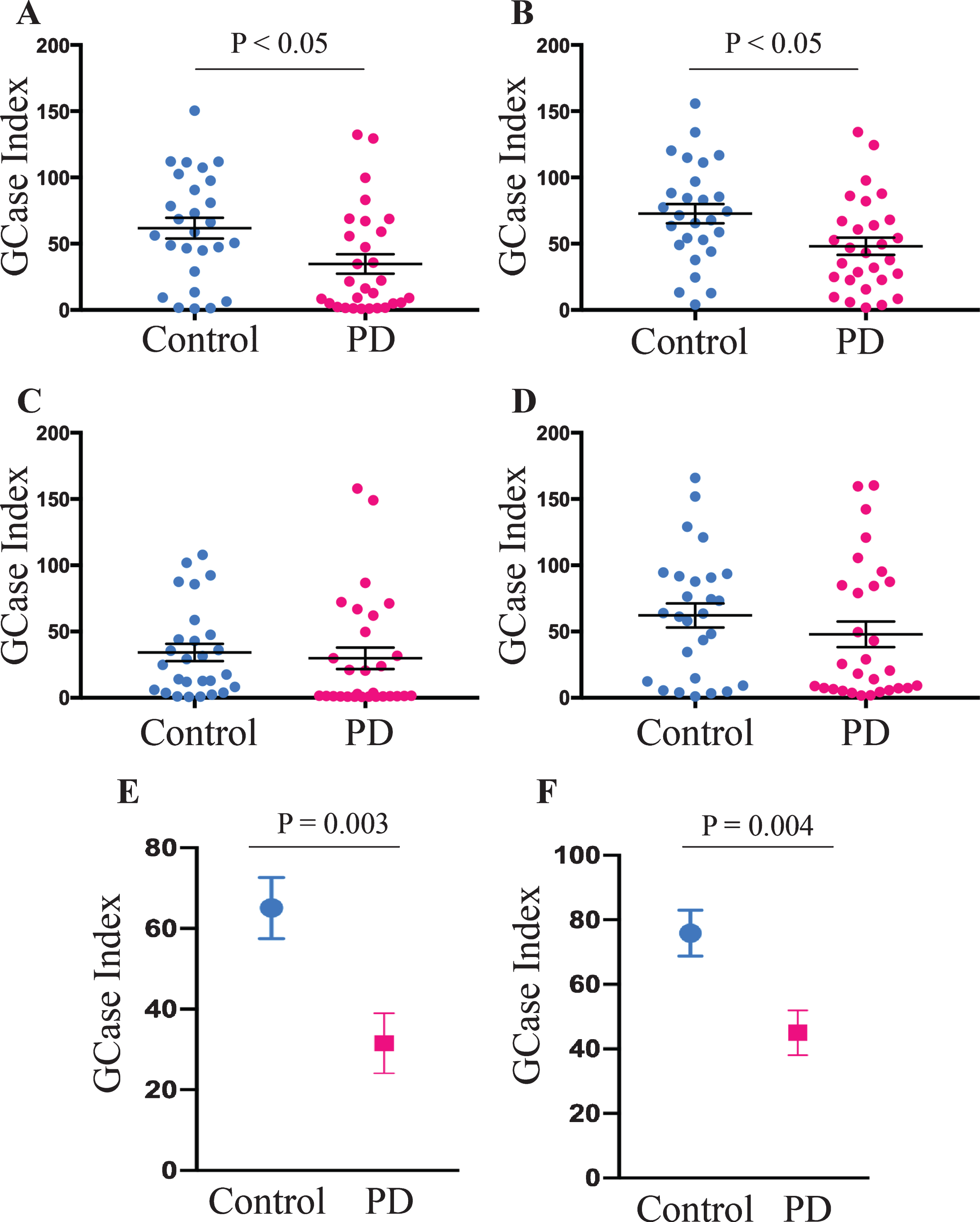
Fig. 3
Reduced GCase activity in intermediate monocytes. GCase activity in classical, intermediate, and non-classical monocyte subsets across the whole cohort (n = 56) was compared using a one-way ANOVA with Tukey’s multiple comparison post hoc test. Data are presented as the mean±SEM with each dot representing an individual participant.
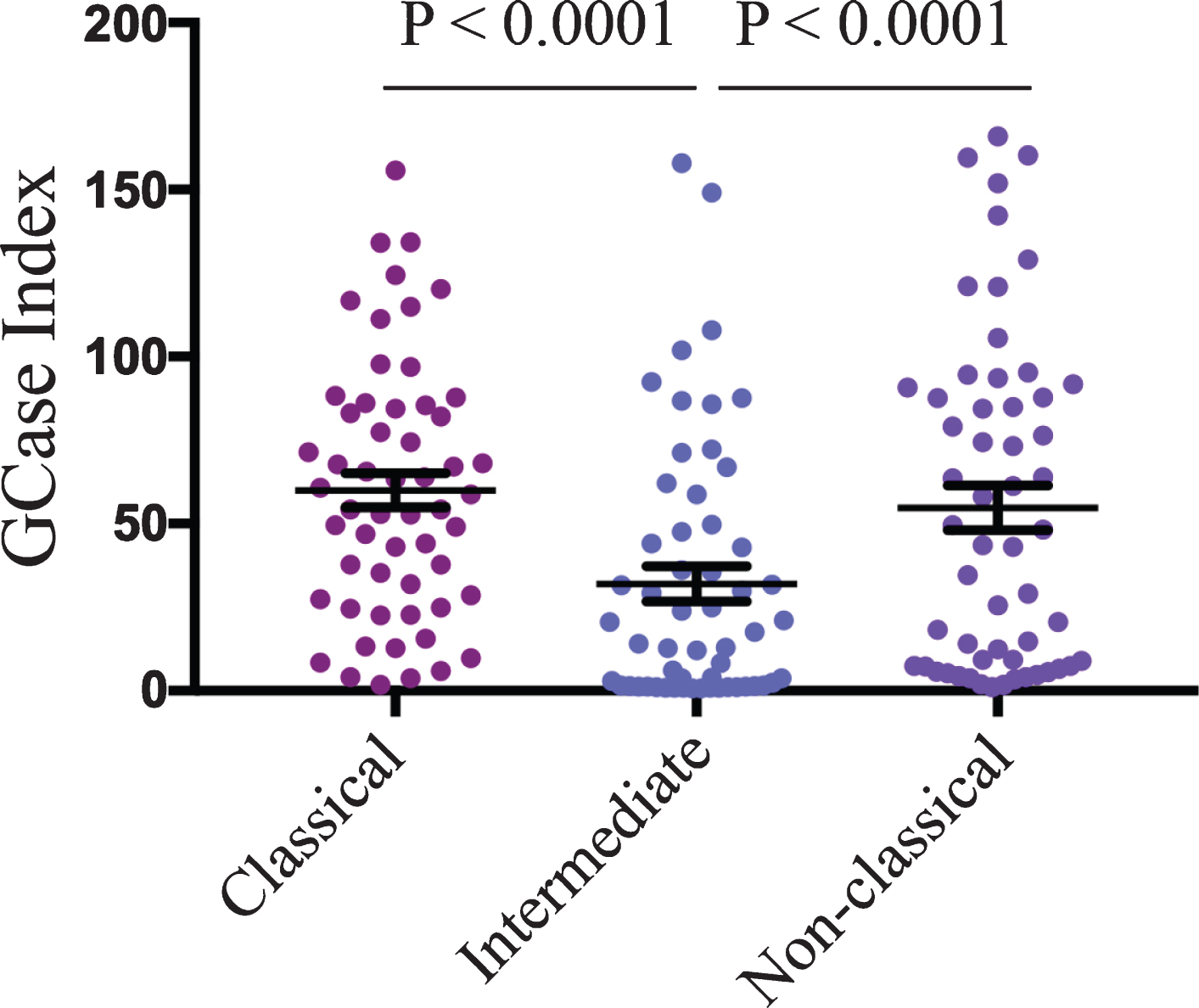
Fig. 4
Classical monocyte GCase activity negatively correlates with PD motor severity. Partial correlation analysis covarying for age, sex, percent of monocytes in PBMCs and storage time in months showed a significant association between classical monocyte GCase activity and participant UPDRSIII score when the whole cohort was included (A) (n = 55), or when just PD patients were included (B) (n = 30).
DISCUSSION
A number of synthetic substrates exist for the measurement of GCase activity, including the commonly used glucosidase substrate 4-methylumbelliferyl β-D-glucopyranoside (4-MUG), which is hydrolyzed to give the quantifiable fluorescent product 4-methylumbelliferone (4-MU) [17, 18], and C12-glucocerebroside, which is hydrolyzed to C12 ceramide and quantified by mass spectrometry [19, 20]. However, the cell permeable quenched substrate PFB-FDGlu, which is hydrolyzed by GCase to release fluorescein, has an advantage over other substrates in that it can be used to directly assess GCase activity in situ in living single cells [21, 22]. Using the PFB-FDGlu substrate in combination with flow cytometry and surface marker antibodies to delineate PBMC types, we have previously shown that GCase activity is specifically reduced in monocytes from PD patients [15]. The current study replicates this finding in a second independent patient cohort, and significantly, was able to confirm this result using cryopreserved PBMCs. That cryopreserved PBMCs can be used for GCase assays can facilitate larger, multi-centre trials utilizing monocyte GCase activity as a biomarker of PD.
In keeping with our previous work, the current study recruited sporadic PD patients that did not have genetic variations in GBA1. Although a number of studies have found reduced GCase activity in brain tissue from sporadic PD patients [6–9], the extent to which peripheral GCase activity may be impaired in sporadic PD is less clear. Studies employing the 4-MUG substrate and whole PBMC lysates have not detected a significant difference in GCase activity between control and sporadic PD patients [23, 24], and while an initial study employing whole blood spots and the C12-glucocerebroside substrate found reduced GCase activity in sporadic PD patients [12], this was not replicated in larger longitudinal studies using the same methodology [13]. Both the 4-MUG probe and blood spot assay assess GCase activity in bulk cell lysates, and outcomes can therefore be affected by the markedly heterogeneous nature of the cells comprising the sample. In contrast, the cell permeable GCase substrate, PFB-FDGlu allows for single cell resolution and specific quantification of in situ GCase activity in distinct cell types. That we found decreased GCase activity in monocytes, and in particular classical monocytes, in sporadic PD patients may suggest a cell type specific effect that is not captured when examining heterogeneous biological samples in which monocytes comprise only a small fraction. Indeed, there is increasing evidence that monocyte dysfunction may contribute to PD [25–27], and a number of PD risk alleles are enriched in monocytes [28, 29]. The inverse correlation observed between monocyte GCase activity and motor severity scores may further indicate a role for monocytes in PD progression; however, larger cohorts and longitudinal assessment are needed to confirm this association.
Our data also indicates that monocyte GCase activity is variable, with a number of PD patients having reduced GCase activity but also a number having levels similar to the matched controls. Therefore, another potential use of monocyte GCase activity measurements may be in the stratification of sporadic PD patients with low GCase activity into clinical trials targeting the GCase enzyme. However, more work is needed to consolidate our findings across larger cohorts and to develop a better understanding of what factors may influence the stability of GCase activity over time. The use of the fluorescent 4-MUG substrate has also been associated with falsely identifying low GCase activity [30]. Therefore, it will also be important to confirm monocyte GCase activity longitudinally and determine how activity relates to disease progression. It will also be important in future studies to include additional orthogonal readouts of GCase activity, such as alterations in levels of glucosylceramide or ceramide species, and/or additional readouts of monocyte lysosomal function. In summary, our results provide a pathway to scalable testing that may have particular relevance for clinical trials, patient phenotyping and/or understanding mechanisms of disease pathogenesis.
ACKNOWLEDGMENTS
This work was jointly funded by the Michael J Fox Foundation for Parkinson’s disease research and the Shake It Up Australia Foundation (#11888.01). GMH holds a NHMRC senior leadership fellowship (#176607). SJGL holds an NHMRC leadership fellowship (#1195830). The Dementia and Movement Disorders Laboratory is supported by ForeFront, a collaborative research group dedicated to the study of non-Alzheimer disease degenerative disorders, funded by NHMRC grants (#1037746, #1095127 and #1132524). We acknowledge Sydney Cytometry for access to flow cytometers and technical assistance. We thank Shikara Keshiya for technical assistance.
CONFLICT OF INTEREST
The authors have no conflicts of interest to report
REFERENCES
[1] | Lesage S , Anheim M , Condroyer C , Pollak P , Durif F , Dupuits C , Viallet F , Lohmann E , Corvol JC , Honore A , Rivaud S , Vidailhet M , Durr A , Brice A , French Parkinson’s Disease Genetics Study Group ((2011) ) Large-scale screening of the Gaucher’s disease-related glucocerebrosidase gene in Europeans with Parkinson’s disease. Hum Mol Genet 20: , 202–210. |
[2] | Blauwendraat C , Reed X , Krohn L , Heilbron K , Bandres-Ciga S , Tan M , Gibbs JR , Hernandez DG , Kumaran R , Langston R , Bonet-Ponce L , Alcalay RN , Hassin-Baer S , Greenbaum L , Iwaki H , Leonard HL , Grenn FP , Ruskey JA , Sabir M , Ahmed S , Makarious MB , Pihlstrom L , Toft M , van Hilten JJ , Marinus J , Schulte C , Brockmann K , Sharma M , Siitonen A , Majamaa K , Eerola-Rautio J , Tienari PJ , andMe Research T, Pantelyat A , Hillis AE , Dawson TM , Rosenthal LS , Albert MS , Resnick SM , Ferrucci L , Morris CM , Pletnikova O , Troncoso J , Grosset D , Lesage S , Corvol JC , Brice A , Noyce AJ , Masliah E , Wood N , Hardy J , Shulman LM , Jankovic J , Shulman JM , Heutink P , Gasser T , Cannon P , Scholz SW , Morris H , Cookson MR , Nalls MA , Gan-Or Z , Singleton AB ((2020) ) Genetic modifiers of risk and age at onset in GBA associated Parkinson’s disease and Lewy body dementia. Brain 143: , 234–248. |
[3] | Gegg ME , Schapira AHV ((2018) ) The role of glucocerebrosidase in Parkinson disease pathogenesis. FEBS J 285: , 3591–3603. |
[4] | Gatto EM , Da Prat G , Etcheverry JL , Drelichman G , Cesarini M ((2019) ) Parkinsonisms and glucocerebrosidase deficiency: A comprehensive review for molecular and cellular mechanism of glucocerebrosidase deficiency. Brain Sci 9: , 30. |
[5] | Blandini F , Cilia R , Cerri S , Pezzoli G , Schapira AHV , Mullin S , Lanciego JL ((2019) ) Glucocerebrosidase mutations and synucleinopathies: Toward a model of precision medicine. Mov Disord 34: , 9–21. |
[6] | Murphy KE , Gysbers AM , Abbott SK , Tayebi N , Kim WS , Sidransky E , Cooper A , Garner B , Halliday GM ((2014) ) Reduced glucocerebrosidase is associated with increased alpha-synuclein in sporadic Parkinson’s disease. Brain 137: , 834–848. |
[7] | Moors TE , Paciotti S , Ingrassia A , Quadri M , Breedveld G , Tasegian A , Chiasserini D , Eusebi P , Duran-Pacheco G , Kremer T , Calabresi P , Bonifati V , Parnetti L , Beccari T , van de Berg WDJ ((2018) ) Characterization of brain lysosomal activities in GBA-related and sporadic Parkinson’s disease and dementia with Lewy bodies. Mol Neurobiol 56: , 1344–1355. |
[8] | Gegg ME , Burke D , Heales SJ , Cooper JM , Hardy J , Wood NW , Schapira AH ((2012) ) Glucocerebrosidase deficiency in substantia nigra of Parkinson disease brains. Ann Neurol 72: , 455–463. |
[9] | Chiasserini D , Paciotti S , Eusebi P , Persichetti E , Tasegian A , Kurzawa-Akanbi M , Chinnery PF , Morris CM , Calabresi P , Parnetti L , Beccari T ((2015) ) Selective loss of glucocerebrosidase activity in sporadic Parkinson’s disease and dementia with Lewy bodies. Mol Neurodegener 10: , 15. |
[10] | Sardi SP , Cedarbaum JM , Brundin P ((2018) ) Targeted therapies for Parkinson’s disease: From genetics to the clinic. Mov Disord 33: , 684–696. |
[11] | Balestrino R , Schapira AHV ((2018) ) Glucocerebrosidase and Parkinson disease: Molecular, clinical, and therapeutic implications. Neuroscientist 24: , 540–559. |
[12] | Alcalay RN , Levy OA , Waters CC , Fahn S , Ford B , Kuo SH , Mazzoni P , Pauciulo MW , Nichols WC , Gan-Or Z , Rouleau GA , Chung WK , Wolf P , Oliva P , Keutzer J , Marder K , Zhang X ((2015) ) Glucocerebrosidase activity in Parkinson’s disease with and without GBA mutations. Brain 138: , 2648–2658. |
[13] | Alcalay RN , Wolf P , Chiang MSR , Helesicova K , Zhang XK , Merchant K , Hutten SJ , Scherzer C , Caspell-Garcia C , Blauwendraat C , Foroud T , Nudelman K , Gan-Or Z , Simuni T , Chahine LM , Levy O , Zheng D , Li G , Sardi SP , Parkinson’s Progression Markers Initiative ((2020) ) Longitudinal measurements of glucocerebrosidase activity in Parkinson’s patients. Ann Clin Transl Neurol 7: , 1816–1830. |
[14] | Huh YE , Chiang MSR , Locascio JJ , Liao Z , Liu G , Choudhury K , Kuras YI , Tuncali I , Videnovic A , Hunt AL , Schwarzschild MA , Hung AY , Herrington TM , Hayes MT , Hyman BT , Wills AM , Gomperts SN , Growdon JH , Sardi SP , Scherzer CR ((2020) ) beta-Glucocerebrosidase activity in GBA-linked Parkinson disease: The type of mutation matters. Neurology 95: , e685–e696. |
[15] | Atashrazm F , Hammond D , Perera G , Dobson-Stone C , Mueller N , Pickford R , Kim WS , Kwok JB , Lewis SJG , Halliday GM , Dzamko N ((2018) ) Reduced glucocerebrosidase activity in monocytes from patients with Parkinson’s disease. Sci Rep 8: , 15446. |
[16] | Postuma RB , Berg D , Stern M , Poewe W , Olanow CW , Oertel W , Obeso J , Marek K , Litvan I , Lang AE , Halliday G , Goetz CG , Gasser T , Dubois B , Chan P , Bloem BR , Adler CH , Deuschl G ((2015) ) MDS clinical diagnostic criteria for Parkinson’s disease. Mov Disord 30: , 1591–1601. |
[17] | Olivova P , Cullen E , Titlow M , Kallwass H , Barranger J , Zhang K , Keutzer J ((2008) ) An improved high-throughput dried blood spot screening method for Gaucher disease. Clin Chim Acta 398: , 163–164. |
[18] | Chamoles NA , Blanco M , Gaggioli D , Casentini C ((2002) ) Gaucher and Niemann-Pick diseases–enzymatic diagnosis in dried blood spots on filter paper: Retrospective diagnoses in newborn-screening cards. Clin Chim Acta 317: , 191–197. |
[19] | Li Y , Scott CR , Chamoles NA , Ghavami A , Pinto BM , Turecek F , Gelb MH ((2004) ) Direct multiplex assay of lysosomal enzymes in dried blood spots for newborn screening. Clin Chem 50: , 1785–1796. |
[20] | Zhang XK , Elbin CS , Turecek F , Scott R , Chuang WL , Keutzer JM , Gelb M ((2010) ) Multiplex lysosomal enzyme activity assay on dried blood spots using tandem mass spectrometry. Methods Mol Biol 603: , 339–350. |
[21] | Lorincz M , Herzenberg LA , Diwu Z , Barranger JA , Kerr WG ((1997) ) Detection and isolation of gene-corrected cells in Gaucher disease via a fluorescence-activated cell sorter assay for lysosomal glucocerebrosidase activity. Blood 89: , 3412–3420. |
[22] | van Es HH , Veldwijk M , Havenga M , Valerio D ((1997) ) A flow cytometric assay for lysosomal glucocerebrosidase. Anal Biochem 247: , 268–271. |
[23] | Kim HJ , Jeon B , Song J , Lee WW , Park H , Shin CW ((2016) ) Leukocyte glucocerebrosidase and beta-hexosaminidase activity in sporadic and genetic Parkinson disease. Parkinsonism Relat Disord 23: , 99–101. |
[24] | Papagiannakis N , Xilouri M , Koros C , Stamelou M , Antonelou R , Maniati M , Papadimitriou D , Moraitou M , Michelakakis H , Stefanis L ((2015) ) Lysosomal alterations in peripheral blood mononuclear cells of Parkinson’s disease patients. Mov Disord 30: , 1830–1834. |
[25] | Grozdanov V , Bliederhaeuser C , Ruf WP , Roth V , Fundel-Clemens K , Zondler L , Brenner D , Martin-Villalba A , Hengerer B , Kassubek J , Ludolph AC , Weishaupt JH , Danzer KM ((2014) ) Inflammatory dysregulation of blood monocytes in Parkinson’s disease patients. Acta Neuropathol 128: , 651–663. |
[26] | Nissen SK , Shrivastava K , Schulte C , Otzen DE , Goldeck D , Berg D , Moller HJ , Maetzler W , Romero-Ramos M ((2019) ) Alterations in blood monocyte functions in Parkinson’s disease. Mov Disord 34: , 1711–1721. |
[27] | Schlachetzki JCM , Prots I , Tao J , Chun HB , Saijo K , Gosselin D , Winner B , Glass CK , Winkler J ((2018) ) A monocyte gene expression signature in the early clinical course of Parkinson’s disease. Sci Rep 8: , 10757. |
[28] | Raj T , Rothamel K , Mostafavi S , Ye C , Lee MN , Replogle JM , Feng T , Lee M , Asinovski N , Frohlich I , Imboywa S , Von Korff A , Okada Y , Patsopoulos NA , Davis S , McCabe C , Paik HI , Srivastava GP , Raychaudhuri S , Hafler DA , Koller D , Regev A , Hacohen N , Mathis D , Benoist C , Stranger BE , De Jager PL ((2014) ) Polarization of the effects of autoimmune and neurodegenerative risk alleles in leukocytes. Science 344: , 519–523. |
[29] | Dzamko N , Geczy CL , Halliday GM ((2014) ) Inflammation is genetically implicated in Parkinson’s disease. Neuroscience 302: , 89–102. |
[30] | Wolf P , Alcalay RN , Liong C , Cullen E , Pauciulo MW , Nichols WC , Gan-Or Z , Chung WK , Faulkner T , Bentis C , Pomponio RJ , Ma X , Kate Zhang X , Keutzer JM , Oliva P ((2018) ) Tandem mass spectrometry assay of beta-glucocerebrosidase activity in dried blood spots eliminates false positives detected in fluorescence assay. Mol Genet Metab 123: , 135–139. |


