
The molecular genetics of Joubert syndrome and related ciliopathies: The challenges of genetic and phenotypic heterogeneity
Abstract
Joubert syndrome (JS; MIM PS213300) is a rare, typically autosomal recessive disorder characterized by cerebellar vermis hypoplasia and a distinctive malformation of the cerebellum and brainstem identified as the “molar tooth sign” on brain MRI. Other universal features include hypotonia with later ataxia and intellectual disability/developmental delay, with additional features consisting of oculomotor apraxia and abnormal respiratory pattern. Notably, other, more variable features include renal cystic disease, typically nephronophthisis, retinal dystrophy, and congenital hepatic fibrosis; skeletal changes such as polydactyly and findings consistent with short-rib skeletal dysplasias are also seen in many subjects. These pleiotropic features are typical of a number of disorders of the primary cilium, and make the identification of causal genes challenging given the significant overlap between JS and other ciliopathy conditions such as nephronophthisis and Meckel, Bardet-Biedl, and COACH syndromes. This review will describe the features of JS, characterize the 35 known genes associated with the condition, and describe some of the genetic conundrums of JS, such as the heterogeneity of founder effects, lack of genotype-phenotype correlations, and role of genetic modifiers. Finally, aspects of JS and related ciliopathies that may pave the way for development of therapeutic interventions, including gene therapy, will be described.
1Introduction
Joubert syndrome (JS; OMIM PS213300) is a predominantly autosomal recessive ciliopathy condition characterized by a distinctive cerebellar and brainstem defect on cranial MRI known as the “molar tooth sign” because of its resemblance to the cross-section of a tooth on axial imaging [64, 65]. Its three key diagnostic criteria have been established as (1) the “molar tooth sign” (MTS), (2) hypotonia in infancy with later ataxia, and (3) developmental delays/intellectual disability. There are several other more variable features, including breathing problems— often episodic tachypnea and/or apnea— especially in infancy, and abnormal eye movements typically characterized by oculomotor apraxia or the inability in initiating voluntary saccades [70]. In addition, a number of findings are sometimes found in those with JS such as retinal disease, cystic kidney disease including nephronophthisis, congenital hepatic fibrosis, and skeletal features such as polydactyly and small ribcage; many of these are also shared by other ciliopathy conditions [6, 118].
Since the first gene for JS, NPHP1, was identified in 2004 [72], over 30 genes have been implicated in its causation [70]. However, most of these genes cause less than 10% of cases each, and for several genes, the assignment is based on only one or a handful of cases. All of the genes identified thus far are localized to or play a role in the function of the sub-cellular structure, the primary cilium, particularly at the transition zone of the cilium where it joins the plasma membrane [36]. The primary cilium is the appendage on almost all cells that functions in the cell’s ability to establish a pattern of left-right symmetry during development, allows it to transduce signals from the environment, and carries out fundamental intracellular processes via signaling pathways such as Sonic Hedgehog and others [82].
This review will focus on the molecular genetics of JS, while also including genetics-based discussion of other relevant ciliary disorders with overlapping phenotypes, and will provide some examples of potential future treatments to capitalize based on what is known about gene function and ciliary biology.
2Diagnostic criteria for Joubert syndrome
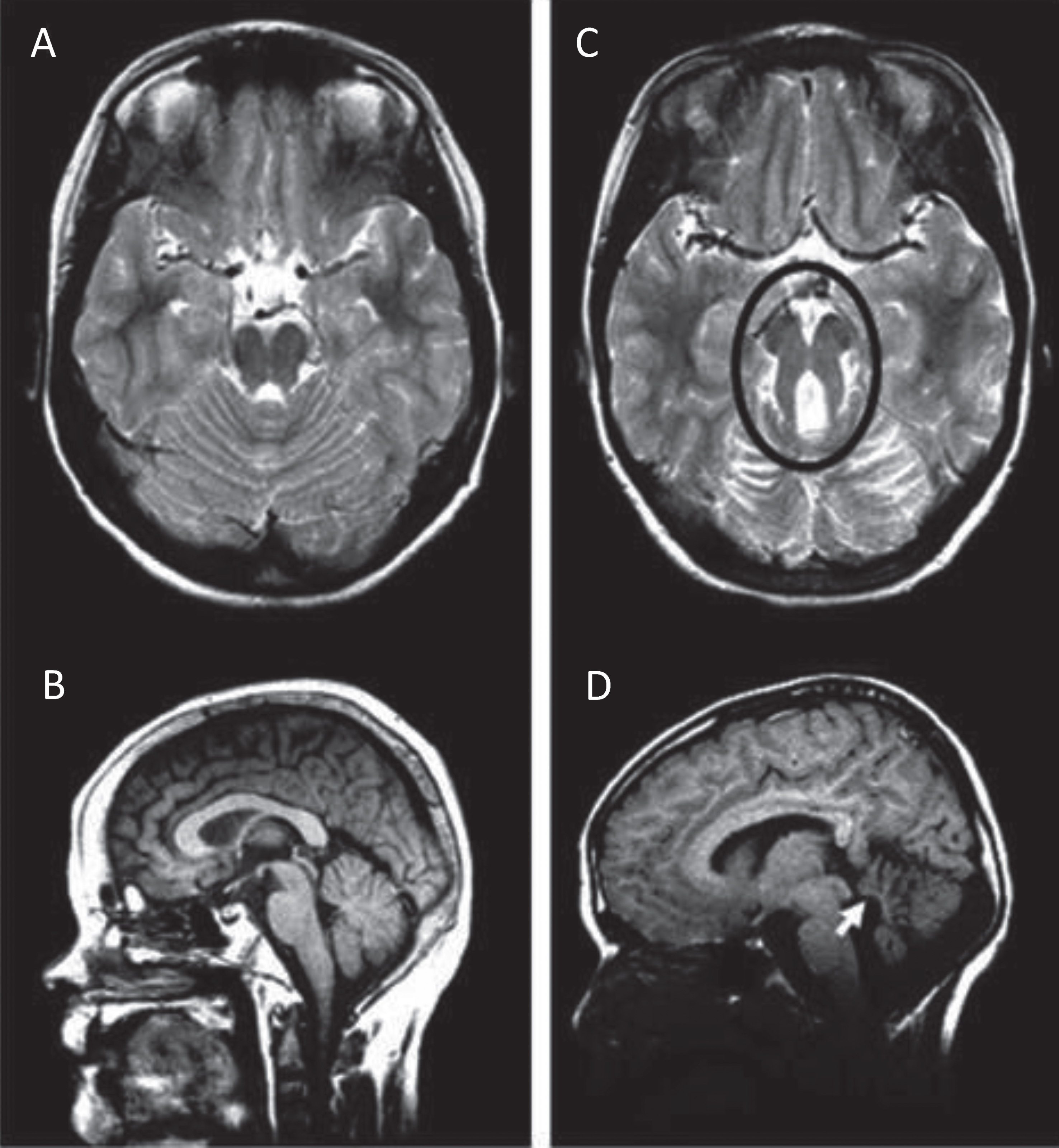
The diagnosis of JS has been established as requiring 3 key elements: the radiologic finding of the “molar tooth sign” (MTS; Fig. 1A, C) accompanied by cerebellar vermis hypoplasia, as well as hypotonia or low muscle tone and developmental delays or intellectual disability. The MTS is visualized on axial views from magnetic resonance imaging (MRI) studies through the junction of the midbrain and pons and consists of the mesencephalon (the “crown” of the tooth), typically with a deepened interpeduncular fossa, and prominent, thickened, and generally straight superior cerebellar peduncles (the “roots” of the tooth) [64, 65]. The MTS is generally associated with hypoplasia of the midline portion of the cerebellum (the cerebellar vermis), a required feature in JS. On sagittal MRI, an enlarged, misshapen and/or rostral shifting of the fastigium of the 4th ventricle is often visualized (Fig. 1B, D). Although a high-resolution MRI scan with 3 mm sections is recommended for visualizing this radiologic sign, the MTS can sometimes be viewed on good quality, high density computed tomography (CT) scans. In reality, a radiologic finding of the MTS is often considered “pathognomonic” for JS, and certainly this discovery in the prenatal or neonatal period would warrant a presumptive diagnosis of JS, as the other required features are likely to become apparent with time (see refs [27, 80] for prenatal radiologic features in JS). In addition, there is a spectrum of MTS findings, with milder versions seen in association with some causative genes [31, 72], and without examination of multiple MRI cuts for subtle findings of the MTS and vermis hypoplasia, even experienced neuroradiologists may miss this diagnostic hallmark.
Fig.1
Molar Tooth Sign in Joubert syndrome. T2-weighted MRI images on axial view (A) and mid-sagittal view (B) through the cerebellum and brainstem of a normal individual showing intact cerebellar vermis. T2-weighted MRI image on axial view (C) and mid-sagittal view (D) through the cerebellum and brainstem of a child with JS. The Molar Tooth Sign is circled in black, with rostral shifting or elevation of the roof (fastigium) of the fourth ventricle and vermis hypoplasia (white arrow).
The hypotonia of infancy often gives way to later ataxia. Developmental delays in motor and cognitive development are universal [76, 105], although rare individuals with JS and normal IQ have been described [76]. Of note, a correlation between degree of cerebellar vermis hypoplasia and cognitive impairment has been seen [77]. In fact, a systematic evaluation of the neuropsychological profiles in 76 participants with JS showed an average Full Scale Intelligence Quotient (FSIQ) = 64±15 and relative strengths in receptive language and verbal comprehension, but challenges with processing speed most likely related to cerebellar deficits [105]. Behavioral surveys suggested more emotional and behavioral problems than in the general population, which may worsen with age, but not as severe as other populations with intellectual disability [105].
Although the 3 cardinal features are necessary for a diagnosis, additional features are often described in infants and children with JS. An abnormal respiratory pattern, particularly prominent in infancy, consists of alternating tachypnea and/or apnea; some neonates have had worrisome bouts of apnea requiring pharmacological intervention [63, 103]. In fact, for those with JS under 5 years of age who were deceased, respiratory failure was the leading cause of death [24]. Eye movements characterized as oculomotor apraxia, or the absence of or defect in controlled, voluntary, and purposeful eye movements resulting in jerky eye movements and head thrusting, as well as nystagmus, strabismus, and ptosis of the eyelids, are often identified in those with JS (Fig. 2A). The spectrum of ocular findings in JS has been prospectively ascertained in one large cohort [16] and the published literature on ocular findings in JS has also been reviewed recently by two groups [16, 119], with general findings of delayed visual development and variable visual acuity [16, 119]. Retinopathy has been reported in nearly 1/3, and ocular coloboma in nearly 30%; however, these rarely occur in the same individual. Optic nerve atrophy was reported in only 10–15% of these cohorts [16, 119].
Fig.2
Clinical features in Joubert syndrome. A. Facial features in a girl with JS/COACH syndrome at 27 months of age showing broad forehead, arched eyebrows, strabismus, eyelid ptosis (on right eye), and open mouth configuration indicating reduced facial tone. B. Oral findings in a child with oral– facial– digital syndrome-like features of JS showing midline upper lip cleft (arrowhead), midline groove of tongue, and bumps of the lower alveolar ridge (arrow). C. Left hand of an infant with JS and postaxial polydactyly (arrow). D. Left foot of an infant with JS and preaxial polysyndactyly and fusion of the hallux. E. View from above of an infant with a small occipital encephalocele with protrusion of the occiput of the skull (arrow). (Facial photograph used with permission of the family.) (Previously published in [73] with permission).
![Clinical features in Joubert syndrome. A. Facial features in a girl with JS/COACH syndrome at 27 months of age showing broad forehead, arched eyebrows, strabismus, eyelid ptosis (on right eye), and open mouth configuration indicating reduced facial tone. B. Oral findings in a child with oral– facial– digital syndrome-like features of JS showing midline upper lip cleft (arrowhead), midline groove of tongue, and bumps of the lower alveolar ridge (arrow). C. Left hand of an infant with JS and postaxial polydactyly (arrow). D. Left foot of an infant with JS and preaxial polysyndactyly and fusion of the hallux. E. View from above of an infant with a small occipital encephalocele with protrusion of the occiput of the skull (arrow). (Facial photograph used with permission of the family.) (Previously published in [73] with permission).](https://content.iospress.com:443/media/trd/2019/4-1-2/trd-4-1-2-trd190041/trd-4-trd190041-g002.jpg)
Phenotypic variability is a key component of JS. There are pleiotropic and highly variable features described in other organ systems that can provide clues to sub-phenotyping, and in some cases, even hints at underlying genetic causes. In addition to the eye findings described above, significant morbidity can arise in those with JS and renal disease (nephronophthisis or cystic kidneys) or congenital hepatic fibrosis. Renal failure is the leading cause of death in those over the age of 5 years [24]. In addition, they can have oral or tongue hamartomas (Fig. 2B), skeletal changes (polydactyly (Fig. 2C, D), syndactyly, and/or narrow thoracic ribcage), and occipital encephalocele (Fig. 2E) [6, 118]. The polydactyly can be postaxial (Fig. 2C), preaxial (Fig. 2D), and occasionally central or mesaxial. Congenital heart defects, cleft palate, skin findings related to those of cranioectodermal dysplasia (sparse hair, small, widely spaced teeth or hypodontia, and/or abnormal nails), situs defects, and obesity are less common but occasionally described. Although ocular colobomas and occipital encephaloceles are developmental defects present at birth, many other features evolve over time and with age, so regular monitoring is necessary to ensure timely diagnosis and treatment if appropriate [70]. Many of these features are typical in other ciliopathy disorders such as Meckel syndrome (MKS) or Oral-Facial-Digital (OFD) syndrome, reinforcing the overlapping phenotypic spectrum of these conditions and confounding attempts at tidy genotype-phenotype correlations [114].
3Clinical subtypes of Joubert syndrome
Given the broad clinical heterogeneity of Joubert syndrome, there are some useful sub-categories that can help in classifying the condition, and sometimes, in predicting which additional manifestations may develop in a given individual [14]. However, even within the same family, siblings with the same causative mutations may have variable phenotypes, illustrating that clinical heterogeneity is the norm in this condition.
3.1Pure or classic JS
In this category, “pure” or “classic” JS has the 3 main diagnostic findings and often includes oculomotor apraxia and/or respiratory abnormalities, and may or may not be associated with polydactyly and other less common manifestations [70]. Many children born with JS are initially considered in this category, until they exhibit manifestations of retinal and/or renal or hepatic disease with age.
3.2JS with retinal disease (JS-Ret)
This form of JS is associated with pigmentary retinopathy that resembles classic retinitis pigmentosa (RP). In some newborns, the most severe manifestations can resemble congenital blindness with attenuated electroretinogram in the spectrum of Leber Congenital Amaurosis (LCA) [16]. The retinal degeneration may develop with age but may not be as rapidly progressive as in some genetic forms of RP [103]. Individuals with retinal disease rarely have ocular colobomas [16]. Overall, retinal dystrophy has been reported in 24–32% of individuals with the condition [16, 28] and has been strongly linked to the AHI1, ARL3, and CEP290 genes [3, 119].
3.3JS with renal disease (JS-Ren)
The kidney disease in JS is generally one of two forms: nephronophthisis (NPHP) or cystic kidneys, although both of these can be part of a spectrum of cystic renal diseases. NPHP manifests as chronic tubulointerstitial nephropathy, and may present in the first or second decade of life with urine-concentrating defects and increased thirst, often progressing to end-stage renal disease (ESRD) within a decade. In juvenile NPHP, the most typical form in JS, ESRD occurs at a median of 13 years [45]. Pathological evaluation of the kidneys reveals corticomedullary cysts, atrophy and interstitial fibrosis; ultrasound evaluation may demonstrate small, scarred kidneys with increased echogenicity at the corticomedullary junction [33, 99]. Another form of kidney disease manifests as enlarged kidney with multiple cysts resembling autosomal recessive polycystic kidney disease, with attendant risk of hypertension [33, 41]. In several surveys, between 23–32% of those with JS have renal manifestations [26, 33, 118].
3.4JS with oculorenal disease (JS-OR)
The combination of kidney disease and retinopathy is often seen in the same person and is associated with a number of genes that cause JS. Historically, the combination of RP and nephronophthisis was known as Senior-Løken syndrome [14, 71] and has been reported in about a third of those with kidney disease [118].
3.5JS with hepatic disease (JS-H)
JS with hepatic disease is rarely symptomatic at birth, although it is characterized by congenital hepatic fibrosis, a developmental disorder of the portobiliary system caused by dysfunctional cilia. The related ciliopathy, Meckel syndrome, also has congenital hepatic fibrosis with ductal plate malformation as a consistent finding [40]. Features include persistence of embryonic ductal plate structures, cystic dilation of intrahepatic biliary ducts, and progressive fibrosis of the portal tracts [104]. Symptoms are generally related to portal hypertension, and individuals can present with elevated liver enzymes, or with advanced disease, recurrent cholangitis and/or gastroesophageal variceal bleeding with thrombocytopenia. Liver disease is often associated with ocular coloboma, a developmental defect of the eye, and can also be associated with kidney disease as well.
3.6JS with oral-facial-digital features (JS-OFD)
Oral features include midline cleft lip and/or palate, midline groove of the tongue, gum or tongue hamartomas, and oral frenulae (Fig. 2B). Other facial features include hypertelorism and micrognathia or recessed jaw. The polydactyly in JS is often postaxial (Fig. 2C) although preaxial polydactyly of the great toes is not uncommon (Fig. 2D); it can be found in any combination on hands and/or feet [70]. Overall, polydactyly has been described in 13–15% of individuals with JS [6, 118]. These features are typical in the genetically heterogeneous group of ciliopathies known as oral-facial-digital (OFD) syndromes. A distinctive form of midline polydactyly known as mesaxial polydactyly and associated with a Y-shaped metacarpal is described in a specific type of OFD known as OFD type VI [12, 37].
3.7JS with acrocallosal features (JS-AC)
In addition to the described structural cerebellar malformations in the human brain in JS, some affected individuals also have abnormalities of the corpus callosum, which connects the two cerebral hemispheres. On careful inspection of MRI scans, one survey found that up to 80% of JS subjects had some sort of callosal dysgenesis [92]. Some families with JS-AC features have mutations in the KIF7 gene, which also causes acrocallosal syndrome with polydactyly and hydrocephalus [6, 79].
3.8JS with Jeune asphyxiating thoracic dystrophy (JS-JATD)
Skeletal abnormalities described in short-rib polydactyly phenotypes include narrow thoracic rib cage, short ribs, shortened tubular bones, and a ‘trident’ appearance of the acetabular roof, with or without polydactyly. Similar skeletal features have been described in subjects with JS and the MTS, most typically, short stature, narrow rib cage, rhizomelic limb shortening, cone-shaped phalangeal epiphyses, brachydactyly, and polydactyly [42, 58, 95].
4Prevalence and epidemiology of Joubert syndrome
The prevalence of JS has been estimated to be between 1:80,000 and 1:100,000 [14, 53, 73], but this may represent an underestimate as more causative genes and private mutations are identified, as well as a broader range of phenotypic findings. JS is a predominantly autosomal recessive disorder first described in relatively isolated groups with high rates of consanguinity, including in French-Canadian, Ashkenazi Jewish, and Arab populations [30, 100–102]. Interestingly, there are multiple founder mutations in several genes for each of these populations. The original French-Canadian family described by Joubert et al in 1969 [49] was known to have distant consanguinity related to a common founder 10 generations removed in France. However, there are at least 3 different genes known to cause JS in families from this region of Canada (CPLANE1, CC2D2A, NPHP1), and the breadth of mutations even within genes linked to an inbred population is highly variable. In fact, many affected individuals who are French-Canadian harbor heterozygous pathogenic variants [100], indicating that the concept of a single “founder” mutation in a particular consanguineous population does not apply to JS. This phenomenon also applies to families from the Middle East, where although there is a high rate of consanguinity, there is a diversity in the number of causative genes [6]. However, in many families of relatively close Arab descent, with first cousin or comparable relatedness, the pathogenic variants are likely to be homozygous, reflecting identity-by-descent. The heterogeneity of causative genes, and in some cases, mutations in an inbred population has also been reported in other ciliopathy disorders. In Newfoundland, for example, multiple different founder effects appear to be causative of Bardet-Biedl syndrome (BBS; [67]). A plausible molecular explanation has not been postulated.
5Genes associated with Joubert syndrome
There are 35 genes implicated as causative for JS (Table 1). Although most are inherited in an autosomal recessive manner, as are the majority of ciliopathies, one, OFD1 (Oral Facial Syndrome 1) demonstrates X-linked recessive inheritance. A few genes have been suggested to have autosomal dominant inheritance, but this is highly speculative as exhaustive screening for other variants may not have been performed. Collectively, these genes account for between 62–94% of cases of JS [6, 75, 118]. The high rate of 94% with identified genetic causes in one survey [118] may be related in part to the large proportion of subjects with hepatic involvement and COACH phenotypes, with better known and characterized genetic causes. Of note, almost all of the genes have also been implicated in other ciliopathy disorders, such as MKS, NPHP, OFD, BBS, COACH syndrome, and others (see column of “Allelic disorders” in Table 1). In general, it is difficult to identify clear-cut genotype-phenotype correlations for many of these genes, but a few distinguishing features are included in the table, as well as genes where founder effects have been identified. In some cases, a more severe or even lethal condition, such as MKS, has been associated with more severe pathogenic changes such as frameshift or nonsense mutations causing loss-of-function rather than milder missense variants often associated with a non-lethal phenotype, such as JS [14]— See Additional Challenges below.
Table 1
Genetic causes of Joubert syndrome, listed in ascending order by chromosomal locus, starting with chromosome 1
| Chromosome | Phenotype in OMIM | Inheritance | Gene/ | Gene/ | Allelic | Sub-cellular | Distinguishing phenotypic | |
| location | pattern | Locus | Locus | Disorders | Localization | features including | ||
| MIM | consanguineous populations1 | |||||||
| number | ||||||||
| 1 | 1p36.32 | Joubert syndrome 25 | AR | CEP104 | 616690 | none | Axoneme tip | French-Canadian; 3 families reported with “Pure” form of JS [100] |
| 2 | 2q13 | Joubert syndrome 4 | AR | NPHP1 | 607100 | NPHP, SLS, BBS | TZ | A common 290 kb deletion accounts for ∼20% of NPHP [72, 99] |
| 3 | 2q33.1 | Joubert syndrome 14 | AR | TMEM237 | 614423 | MKS | TZ | Hutterite; cystic renal disease and HTN [47] |
| 4 | 2q37.1 | Joubert syndrome 30 | AR | ARMC9 | 617612 | none | BB | Pure JS or with RP, PD [116] |
| 5 | 2q37.1 | Joubert syndrome 22 | AR | PDE6D | 602676 | none | Lipidated protein transport | Renal hypoplasia, retinal dystrophy, PD [109] |
| 6 | 3q11.1-q11.2 | Joubert syndrome 8 | AR | ARL13B | 608922 | none | Lipidated protein transport | “Pure” JS or RP plus ENC or obesity [17, 107] |
| 7 | 4p15.32 | Joubert syndrome 9 | AR | CC2D2A | 612013 | MKS, COACH | TZ; BB appendages | 8–11% of JS cases; French-Canadian [7, 39, 68, 100] |
| 8 | 5p13.2 | Joubert syndrome 17 | AR | CPLANE1 (previously C5ORF42) | 614571 | MKS, OFD | TZ | 8–14% of JS; OFD VI features; Dutch; French-Canadian [60, 100, 102] |
| 9 | 5q23.2 | Joubert syndrome 31 | AR | CEP120 | 613446 | JATD, MKS, OFD | BB-centrosome | Skeletal dysplasia phenotype in some [84, 95] |
| 10 | 6q23.3 | Joubert syndrome 3 | AR | AHI1 | 608894 | none | TZ, BB, centriole | 7–10% of JS; RP often and sometimes renal [25, 74, 113] |
| 11 | 7q32.2 | Joubert syndrome 15 | AR | CEP41 | 610523 | MKS | BB, axoneme | PD; occasional RP [56] |
| 12 | 8q13.1-q13.2 | Joubert syndrome 21 | AR | CSPP1 | 611654 | MKS, JATD | BB | 2–4% of JS; Hutterite; skeletal dysplasia in some [1, 96, 112] |
| 13 | 8q22.1 | Joubert syndrome 6 | AR | TMEM67 (previously MKS3) | 609884 | MKS, BBS, COACH, NPHP | TZ | 6–20% of JS; renal disease; COACH features [5, 15, 118] |
| 14 | 9q34.3 | Joubert syndrome 1 | AR | INPP5E | 613037 | MORM | Axoneme-signaling | 2–4% of JS; obesity and micropenis [11, 48] |
| 15 | 10q24.1 | Joubert syndrome 18 | AR | TCTN3 | 613847 | OFD IV, MKS | TZ | Skeletal dysplasia [108] |
| 16 | 10q24.32 | Joubert syndrome 32 | AR | SUFU | 607035 | Medullo-blastoma, BCNS2 | Ciliary tip | PD, PMG [22] |
| 17 | 10q24.32 | Joubert syndrome 35 | AR | ARL3 | 604695 | RP | Lipidated protein transport | RP, kidney cysts [3] |
| 18 | 11q12.2 | Joubert syndrome 16 | AR | TMEM138 | 614459 | MKS, OFD | TZ | Coloboma, RP, cystic kidney or NPHP, PD, ENC [57] |
| 19 | 11q12.2 | Joubert syndrome 2 | AR | TMEM216 | 613277 | MKS, OFD VI | TZ | 2-3% of JS; Ashkenazi Jewish [30, 114] |
| 20 | 12q21.32 | Joubert syndrome 5 | AR | CEP290 | 610142 | MKS, BBS, LCA, SLS | TZ, centriolar satellite | 7–10% of JS; 20% of LCA; broad range of findings; Japanese [90, 106, 115] |
| 21 | 12q21.33 | Joubert syndrome3 | AR | POC1B | 615973 | LCA, CRD | BB | RP/LCA, large polycystic kidneys [10] |
| 22 | 12q24.11 | Joubert syndrome 13 | AR | TCTN1 | 609863 | none | TZ, BB | Pachygyria [35] |
| 23 | 12q24.31 | Joubert syndrome 24 | AR | TCTN2 | 613846 | MKS, NPHP | TZ, BB, axoneme | NPHP, liver fibrosis [87, 94] |
| 24 | 13q21.3-q22.1 | Joubert syndrome 33 | AR | PIBF1 | 607532 | JATD, RP | BB-centrosome | Skeletal dysplasia, RP [120] |
| 25 | 14q23.1 | Joubert syndrome 23 | AR | KIAA0586 | 610178 | Hydrolethalus, JATD | BB | 2–7% of JS [2, 9, 62] |
| 26 | 15q26.1 | Joubert syndrome 12 | AR | KIF7 | 611254 | Hydrolethalus, ACS | Axoneme (tip) | Macrocephaly, ACC, PD [21, 79] |
| 27 | 16p12.1 | Joubert syndrome 26 | AR | KATNIP (previously KIAA0556) | 616650 | None | BB | “Pure” JS; PIT [85, 86] |
| 28 | 16q12.1 | Joubert syndrome 19 | AD, AR | ZNF423 | 604557 | NPHP | nucleus | NPHP, situs inversus [18] |
| 29 | 16q12.2 | Joubert syndrome 7 | AR | RPGRIP1L | 610937 | MKS, COACH | TZ, BB, centrosome | 1–4% of JS; broad range of findings; RP [23, 28, 52, 121] |
| 30 | 16q23.1 | Joubert syndrome 20 | AR | TMEM231 | 614949 | MKS, OFD | TZ | Renal cysts, RP, and PD; French-Canadian [61, 100, 101] |
| 31 | 17p13.1 | Joubert syndrome 29 | AR | TMEM107 | 616183 | MKS, OFD VI | TZ | RP, PD [55, 93] |
| 32 | 17p11.2 | Joubert syndrome 27 | AR | B9D1 | 614144 | MKS | TZ | “Pure” JS [83] |
| 33 | 17q22 | Joubert syndrome 28 | AR | MKS1 | 609883 | MKS, BBS | TZ, centrosome | 2–6% of JS [51, 83, 98] |
| 34 | 19q13.2 | Joubert syndrome 34 | AR | B9D2 | 611951 | MKS | TZ | PD, ENC, CP [6] |
| 35 | Xp22.2 | Joubert syndrome 10 | XLR | OFD1 | 300170 | OFD I (AD), RP | BB distal end, centriolar satellite | ENC, hydrocephalus, PMG, macrocephaly, PD, RP, renal cysts [20, 32] |
General references for these tables include [38, 70, 82] and OMIM. 1For those genes that account for >1% of JS cases in several large surveys ([6, 118]), the frequency is noted. 2Dominant loss-of-function mutations in the SUFU tumor suppressor gene are associated with medulloblastoma, Basal Cell Nevus Syndrome, and susceptibility to familial meningioma. 3Pathogenic variants in this gene have been identified in conjunction with the MTS, but this gene has not been formally assigned an OMIM designation in the JS phenotypic series PS213300. This status also applies to the following genes: C2CD3, HYLS1, KIAA0753, IFT80, and CELSR2. Abbreviations: ACC, agenesis of the corpus callosum; ACS, Acrocallosal syndrome; AD, autosomal dominant; AR, autosomal recessive; BB, Basal body; BBS, Bardet-Biedl syndrome; BCNS, Basal Cell Nevus Syndrome; COACH, Colobomas, cognitive impairment (“Oligophrenia”), Ataxia, Cerebellar vermis hypoplasia, and Hepatic fibrosis syndrome; CP, cleft palate; CRD, Cone-rod dystrophy; ENC, encephalocele; HTN, hypertension; JATD, Jeune asphyxiating thoracic dystrophy; JS, Joubert syndrome; LCA, Leber Congenital Amaurosis; MKS, Meckel syndrome; MORM, Mental retardation, Obesity, Retinal dystrophy, Micropenis syndrome; NPHP, nephronophthisis; OFD, oral-facial-digital syndrome; PD, polydactyly; PIT, pituitary abnormalities, including hypopituitarism, micropenis; PMG, polymicrogyria; RP, retinitis pigmentosa; SLS, Senior-Løken syndrome; TZ, transition zone; XLR, X-linked recessive.
6Proteins of the cilium
Ciliopathy conditions are human disorders characterized by dysfunction of either the motile or immotile cilium, a subcellular organelle important for embryonic development, adult homeostasis, and in sensory and/or signaling processes [38]. JS is a ciliopathy disorder of the primary, non-motile cilium found on almost all vertebrate cells [82]. Most of the genes implicated in JS and related ciliopathies localize to either the primary cilium, the basal body and/or the mother centriole from which it arises, or the regulatory proteins and transcription factors that guide its development and function [82]. The primary cilium has been described as an antennae or appendage protruding from the plasma membrane that plays a role in sensing the environment and responding to localized signals [38]. It develops from the docking of a mother centriole via “distal appendages” or transition fibers that link the centriole to the plasma membrane; this is where the centriole extends the 9 axonemal microtubule doublets into the cilium (Fig. 3A, B). The structure at the base of the cilium and junction of plasma membrane and ciliary membrane is known as the transition zone [36]. This “transition zone,” which, along with the transition fibers and Y-fibers, comprise the “ciliary gate,” lies between the basal body and the ciliary axoneme and controls the entry and exit of proteins and lipids into and out of the cilium. Intraflagellar transport (IFT) of this cargo along the microtubules is driven by kinesin (anterograde) and dynein (retrograde) motors and associated IFT proteins and BBS proteins, with lipidated protein transport also playing a role [82].
Fig.3
Diagram of the structure of the Primary Cilium. A. Schematic of the components of a non-motile (primary cilium) showing the axoneme (ciliary stalk), comprised of 9 pairs of microtubules. The cilium extends from a basal body, which derives from the mother centriole. Transition fibers tether the basal body to the base of the ciliary membrane. The transition zone (TZ) is adjacent to the basal body and serves as a gate for passage of proteins and lipids into and out of the cilium. Anterograde transport is accomplished by kinesin motors along with associated proteins and BBsomes propelling cargo along the microtubules; retrograde transport utilizes dynein motors rather than kinesins. Signaling proteins on the ciliary membrane represent the components of the Sonic Hedgehog (SHH) or other receptor complexes. B. Cross-section of the cilium demonstrates the typical 9+0 pattern of doublet microtubes in the axoneme (upper diagram) and the addition of Y-links that connect the doublet microtubes to the ciliary membrane and help form the barrier within the transition zone (bottom diagram). (Derived from [38, 66, 82]).
![Diagram of the structure of the Primary Cilium. A. Schematic of the components of a non-motile (primary cilium) showing the axoneme (ciliary stalk), comprised of 9 pairs of microtubules. The cilium extends from a basal body, which derives from the mother centriole. Transition fibers tether the basal body to the base of the ciliary membrane. The transition zone (TZ) is adjacent to the basal body and serves as a gate for passage of proteins and lipids into and out of the cilium. Anterograde transport is accomplished by kinesin motors along with associated proteins and BBsomes propelling cargo along the microtubules; retrograde transport utilizes dynein motors rather than kinesins. Signaling proteins on the ciliary membrane represent the components of the Sonic Hedgehog (SHH) or other receptor complexes. B. Cross-section of the cilium demonstrates the typical 9+0 pattern of doublet microtubes in the axoneme (upper diagram) and the addition of Y-links that connect the doublet microtubes to the ciliary membrane and help form the barrier within the transition zone (bottom diagram). (Derived from [38, 66, 82]).](https://content.iospress.com:443/media/trd/2019/4-1-2/trd-4-1-2-trd190041/trd-4-trd190041-g003.jpg)
The transition zone appears to be a hotspot for human disease genes associated with ciliopathies. In fact, there are either 2 or 3 main protein complexes, mapped by interactome studies, that form the transition zone [19, 35]: the MKS complex, which includes the Tectonic proteins, B9 domain proteins, coiled-coil proteins such as CC2D2A and CEP290, AHI1, and 5–7 transmembrane or TMEM proteins; and the NPHP complex, including the NPHP proteins, RPGRIP1 L, and others [36]. Linking these 2 domains are CEP290 and NPHP5, sometimes separated out as a distinctive protein domain [38]. These subdivisions are significant, as almost all of the proteins in the MKS complex are encoded by genes implicated in MKS and JS, while the NPHP complex is associated with nephronophthisis-causing genes predominantly [38]. The developmental function of these proteins is just now being discovered and includes roles in Sonic Hedgehog (SHH) signaling, planar cell polarity, cAMP signaling, mTOR pathway activity, and DNA Damage Response (DDR) signaling pathways [66, 99]. In spite of our ever-evolving understanding of ciliary biology, the precise pathological processes that underlie the specific manifestations of the ciliopathies remain elusive.
7Overlap with other ciliopathies: Allelic Disorders
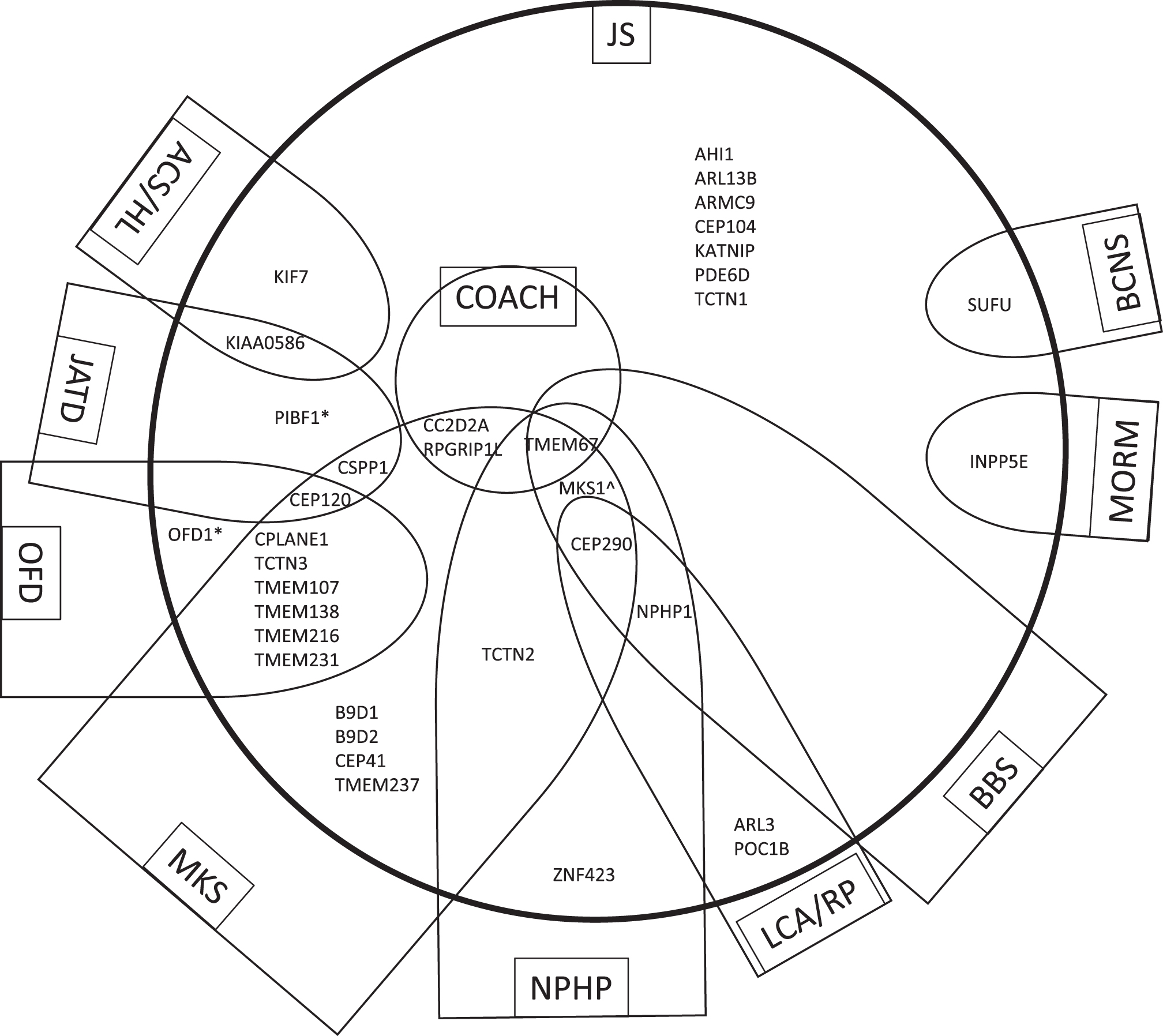
JS is one of several disorders of the primary cilium that have multi-systemic effects, often including involvement of the brain, eyes, kidney, liver, and skeleton. These ciliopathies also overlap in their manifestations, and in their causative genes [66]. The allelic ciliopathy disorders associated with each of the 35 JS-associated genes based on overlapping phenotypes are shown in Fig. 4. The following sub-sections will describe the most common allelic disorders to JS identified thus far:
Fig.4
Genes that cause JS have overlapping phenotypes with other ciliopathies. The 35 genes that cause Joubert syndrome (JS) and are listed in Table 1 are distributed within the large circle in relationship to allelic ciliopathy disorders indicated within each semi-oval (or circle in the case of COACH). The name of each ciliopathy disorder is indicated within the box. Other genes that cause the ciliopathy condition but are not associated with JS are omitted in order to simplify the diagram. These relationships are not fixed, as the phenotypic spectrum associated with ciliopathy genes is constantly evolving. *Can also be associated with RP alone ∧The MKS1 gene has not been described in NPHP alone but this could not be indicated due to constraints of the diagram. Abbreviations: ACS, Acrocallosal syndrome; BBS, Bardet-Biedl syndrome; BCNS, Basal Cell Nevus Syndrome; COACH, Colobomas, cognitive impairment (“Oligophrenia”), Ataxia, Cerebellar vermis hypoplasia, and Hepatic fibrosis syndrome; HL, Hydrolethalus syndrome; JATD, Jeune asphyxiating thoracic dystrophy and related skeletal dysplasias; JS, Joubert syndrome; LCA, Leber Congenital Amaurosis; MKS, Meckel syndrome; MORM, Mental retardation, Obesity, Retinal dystrophy, Micropenis syndrome; NPHP, nephronophthisis; OFD, oral-facial-digital syndrome; RP, retinitis pigmentosa.
7.1Acrocallosal syndrome (ACLS; OMIM 200990)
Acrocallosal syndrome (ACLS; OMIM 200990) is a recessive condition with macrocephaly, agenesis of the corpus callosum, other brain anomalies, intellectual disability, and polydactyly. The previously reported association in some families with KIF7 mutations and either an ACLS, JS or Hydrolethalus syndrome (OMIM PS236680; generally lethal condition with midline brain anomalies such as hydrocephalus or anencephaly) phenotype suggests that they are allelic conditions [79]. The JS gene, KIAA0586 has also been implicated in Hydrolethalus syndrome [2].
7.3Bardet-Biedl syndrome (BBS, OMIM PS209900)
Bardet-Biedl syndrome (BBS, OMIM PS209900) is generally autosomal recessive in inheritance, with some examples of digenic inheritance, and is characterized by cone-rod retinal dystrophy, obesity, postaxial polydactyly, intellectual disability, variable kidney disease, and hypogonadism/genital anomalies. Progressive renal and retinal disease can be a source of morbidity and mortality; at least 19 genes have been implicated thus far [34].
7.4COACH syndrome (COACH, OMIM 216360)
COACH syndrome (COACH, OMIM 216360) the combination of Colobomas, cognitive impairment (“Oligophrenia”), Ataxia, Cerebellar vermis hypoplasia (generally, the MTS), and Hepatic fibrosis [28, 88, 117]. Mutations in the genes TMEM67, CC2D2A, and RPGRIP1 L have been strongly correlated with liver involvement and colobomas [118]. In one large cohort of subjects with JS, 18% had congenital hepatic fibrosis [26]; in another cohort enriched for COACH syndrome, there were 41% with liver involvement [118]. Although renal cystic disease is common in COACH, retinopathy is rarely reported [104].
7.5Jeune asphyxiating thoracic dystrophy (JATD)
Jeune asphyxiating thoracic dystrophy (JATD) is one of a group of autosomal recessive skeletal dysplasias known as short-rib thoracic dystrophies characterized by short stature, a long, narrow thorax with short ribs, short limbs, variable polydactyly, and cystic renal disease. It is well known as a ciliary disease with multiple causative genes, many associated with intraflagellar transport in the cilium. Overlapping genes associated with JS include CEP120, CSPP1, KIAA0586, and PIBF1. In addition to JATD, skeletal features seen in JS are often described in related and overlapping skeletal dysplasias including Ellis-van Creveld syndrome, Mainzer-Saldino syndrome, Short rib-polydactyly syndrome, and Cranioectodermal dysplasia (Sensenbrenner syndrome) [4].
7.6Leber Congenital Amaurosis (LCA, OMIM PS204000)
Leber Congenital Amaurosis (LCA, OMIM PS204000) is a severe form of congenital retinal blindness and nonrecordable electroretinograms. Genetically heterogeneous, with at least 19 genes and typically autosomal recessive but occasionally autosomal dominant inheritance [54]; about 25% of children also have cognitive impairment. The CEP290 gene has been implicated in 20% of LCA due to a cryptic intronic splice donor site as well as JS, MKS, and Senior-Løken syndrome [13, 54].
7.7Meckel syndrome (MKS, OMIM PS249000)
Meckel syndrome (MKS, OMIM PS249000), also known as Meckel-Gruber syndrome, is an autosomal recessive disorder characterized by the triad of cystic dysplastic kidneys, brain malformations of the posterior fossa (usually occipital encephalocele), and the hepatic ductal plate malformation resulting in congenital hepatic fibrosis and proliferation of bile ducts. This condition is often lethal in the prenatal or perinatal periods [43]. At least 21 different genes have been implicated in Meckel syndrome, and for at least 18 of them, pathogenic changes have also been identified in JS (see Table 1). In some cases, more severe variants are associated with Meckel syndrome while variants predicted to have milder effects on protein function are associated with JS [83, 98]. There are also families in which the identical pathogenic variants in the same family can cause both Meckel syndrome and JS [82, 114].
7.8Nephronophthisis (NPHP, OMIM PS256100)
Nephronophthisis (NPHP, OMIM PS256100) was one of the first ciliary disorders to be described, and is clinically manifest as polydipsia and polyuria due to impaired urinary concentrating ability, accompanied by chronic tubulointerstitial nephritis and later development of medullary cysts; it progresses to end-stage renal disease by age 30. About 80–85% of individuals have purely kidney disease, and ∼20% of subjects with juvenile nephronophthisis have a large, homozygous 290-kb deletion in the NPHP1 gene [44, 89, 99]. A few individuals with JS have harbored this same deletion or other mutations in this gene [73]. NPHP is often associated with retinal dystrophy, a combination known as Senior-Løken syndrome (OMIM PS266900). As both NPHP and retinal dystrophy are found in JS, the overlap is considerable.
7.9Oral-facial-digital syndromes (OFD, multiple OMIM)
Oral-facial-digital syndromes (OFD, multiple OMIM) describe a number of disorders with characteristic facial features, oral abnormalities such as tongue lobulations, missing teeth, and extra oral frenula, and polydactyly. At least 13 clinical subtypes have been described, and the spectrum of findings overlaps considerably with MKS, short-rib thoracic dystrophies, and JS. In contrast to other forms of OFD which adhere to autosomal recessive inheritance, OFD1 (OMIM 311200) is an X-linked dominant condition impacting mostly females and causing polycystic kidney disease [110]. The OFD1 gene has also been implicated in a few males with JS and OFD features [32]. OFD IV (Mohr-Majewski syndrome, OMIM 258860) has a spectrum of digital anomalies, brain malformations, tibial dysplasia, short ribs and cystic kidneys with overlap with Meckel and JS; pathogenic variants in TCTN3 have been identified [108]. OFD VI (Varadi-Papp syndrome, OMIM 277170) is also worth mention, as this form of OFD has distinctive midline or mesaxial polydactyly, often with a Y-shaped metacarpal between the middle digits in addition to the facial feature (see Fig. 2B). Mutations in CPLANE1 have been identified in subjects with OFD VI and JS, and there are no clear genotype-phenotype correlations that predict the phenotype (mild JS vs more severe or lethal OFD VI) relative to the severity of the pathogenic variants; in fact, biallelic truncating mutations were just as likely to result in mild JS features as more severe or lethal OFD VI phenotypes [12].
8Additional challenges in genetic diagnoses of ciliopathies
There are several major conundrums in understanding the genetic basis of ciliary disorders. One is the sheer number of genes that is causative for each clinically-defined disorder. Even a seemingly simplified ciliopathy such as nephronophthisis (NPHP) can be accounted for by over 20 genes, all of which are associated with the cilia, centrosome, or mitotic spindle [99]. In fact, most surveys report that about 15–20% of individuals with nephronophthisis are reported to have additional non-renal ciliopathy symptoms, the most common being RP. The NPHP1 gene only accounts for ∼20% of individuals with NPHP, with a homozygous deletion of the entire gene being the major mutation. Even in the situation where an individual harbors this clear biallelic loss-of-function deletion, however, the heterogeneity of the phenotype can range from NPHP to Senior-Løken to JS. Each of the remaining 20+ causative genes accounts for less than 1% of cases of NPHP, suggesting that over 60% of cases do not yet have a known genetic cause, in spite of 20 years of genetic studies on this condition [99]. It is notable that many ciliopathy conditions have multiple causative genes, with few individual genes causing more than 10–20% of cases within a specific condition. This lack of predictive ability and paucity of genotype-phenotype correlations are challenging.
The second conundrum is the extreme genetic heterogeneity of molecular causes and, in particular, the overlap of many causative genes with multiple different ciliopathy conditions. As noted, many of the genes that cause JS can also cause MKS, NPHP, OFD, BBS, and/or Jeune syndrome, or even more obscure ciliopathy conditions. When looking for genotype-phenotype correlations that might explain why the same gene can cause a lethal MKS phenotype as well as a milder JS phenotype and even more focal nephronophthisis, the severity of mutation has been implicated (e.g., 2 mutations in CC2D2A or TMEM67 resulting in frameshift or stop codons cause MKS while “milder” missense mutations cause JS) [8, 68]. Similarly, mutations in TMEM231 can cause MKS, OFD, and JS, with different presentations due to the same mutation even within the same family [61, 101]. In addition, different missense mutations in the same gene can cause disparate phenotypes, such as mutations in CEP290 that can cause JS, BBS, LCA, MKS, or Senior-Løken syndrome; it is likely that alterations in this protein, which straddles the MKS and NPHP modules within the transition zone, have different effects depending on their impact on interacting protein partners or different functional domains of the protein [90, 115]. Another possible explanation for this variability in the different outcomes for pathogenic variants in the same gene is that they may impact discrete isoforms of the protein that have distinctive functions. This is certainly the case for at least one BBS gene, ARL6, where one isoform is part of the BBSome and results in the BBS phenotype, whereas a mutation in a longer isoform has a specific role in photoreceptor maintenance [78]. Finally, it is likely that modifier genes or other epistatic effects such as differences in genetic background play a role. In these situations, proteins with overlapping functions and roles can be revealed. The specific manifestations of the pleiotropic effects of BBS may be partially explained by the unique roles of proteins associated with intraflagellar transport that underlie this condition: for example, ciliary biogenesis is dependent on Hedgehog signaling and is necessary for embryonic development, particularly of the nervous system and limb bud— consequently, mutations in many IFT components are associated with intellectual disability and skeletal dysplasias such as JATD [91]. Likewise, mutations in genes that impact the tissue-specific cargo of the IFT system may result in specific manifestations of ciliopathies [82]. The condition with the most overlap with JS is MKS, and both are associated with causal mutations in overlapping genes whose protein products localize to the transition zone. There is redundancy in the system, as the MKS, NPHP, and BBSome complexes likely have partially overlapping roles in promoting the ciliary localization of proteins and forming a barrier for protein transit into and out of the cilium. For proteins within the transition zone, a non-pathogenic variant may thus modify the phenotypes caused by pathogenic variants in a gene encoding a protein with overlapping or partially redundant function [82].
The third conundrum is the finding of occasional oligogenicity (variants in 2 or more different genes cause the condition; [46]), digenic inheritance, or triallelism in JS, as well as other ciliopathies. Triallelic inheritance refers to the observation that biallelic mutations in one gene are required along with a heterozygous pathogenic variant in a different ciliary gene to cause the disease [50]. Of course, these rare reports may reflect extremes of genetic modifiers, in which variants in other genes in addition to biallelic pathogenic variants in one known causative gene, affects the spectrum and severity of the phenotypic findings [75]. Several specific examples have been described, such as a heterozygous AHI1 mutation in combination with biallelic NPHP1 mutations resulting in a more severe brain phenotype in NPHP [111]. An NPHP1 mutation or deletion may also contribute to the severity of the BBS phenotype caused by 2 BBS biallelic mutations [59]. This has also been demonstrated with a common variant in RPGRIP1 L that modifies the retinal degeneration phenotype in a number of ciliary disorders [52]. Before the advent of next-generation sequencing capabilities, identifying genetic modifiers was challenging, and even targeted gene panels might fail to provide a comprehensive evaluation of all likely contributing modifying genes. One recent effort sought to tease out the prevalence of digenic inheritance, triallelism, and genetic modifiers in a large JS cohort [75]. Although rare disease variants were identified in over 1/3 of affected individuals in addition to the causal pathogenic mutations, they did not correlate with severity of the condition, nor did they support the presence of triallelic inheritance. Importantly, the rate of discordant phenotypes (∼60%) between affected siblings with shared pathogenic variants in a known JS gene suggested that modifying genes play an important role in JS, but they may be difficult to identify [75]. Another comprehensive study of the genetic contributions to a large cohort of subjects with ciliopathy phenotypes in a mostly consanguineous population demonstrated that ciliopathies are, for the most part, Mendelian disorders, as the burden of rare variants in known ciliopathy genes in the cohort did not differ significantly from the rate in a control cohort with non-JS intellectual disability; these authors suggested that stochastic events rather than modifier alleles contributed to phenotypic variability within their population [97].
9Future directions in Joubert syndrome biology, genetics and therapeutics
The sheer number of causative genes for ciliary disorders such as JS can seem overwhelming, and clearly, identifying the causative gene and variant for a given family can be daunting. This is compounded by the lack of clear and consistent founder effects in isolated populations with these conditions. More research to identify additional causative genes will be critical for this effort, and new methodologies such as whole exome and whole genome sequencing are clearly paving the way, although interpretation of the meaning of variants can be quite challenging given the heterogeneity of genotypes and lack of correlation with phenotypes. Certainly, these features point to some level of redundancy in the complex biology of the cilium. And the extreme pleiotropy of organ system involvement in JS and other ciliopathies suggests that a deeper understanding of the roles of ciliary proteins, their interactions, and tissue specificity is necessary. In addition, the emerging role of modifying genes, many of which are ciliary genes as well, points to the role of ciliopathies serving as a bridge between strictly mendelian disorders and more complex polygenic disorders. For the extreme and rare cases of digenic or triallelic inheritance, the concept of threshold for mutational load appears operative. And in general, many of the principles that hold for one ciliary disorder are likely to be generalizable to other ciliopathies, suggesting that there may be some concepts of ciliary genetics and biology that are applicable to several, if not most, ciliopathies. For example, although many genes and several different signaling pathways are implicated in the pathogenesis of NPHP, the different mechanistic processes converge on the final common endpoints of renal tubular damage and fibrosis leading to ESRD [99].
In spite of lack of knowledge about the functional and developmental roles of many ciliarry proteins, there remains hope for development of therapies to impact specific ciliary phenotypes. Rescuing the developmental brainstem and cerebellar malformations that underlie the MTS may not be achievable, at least in the foreseeable future, but interventions to ameliorate the co-occurring morbidities such as retinal, renal, and/or liver disease could significantly impact quality of life for those with JS. One strategy to approach the cystic renal disease of NPHP has been to treat the elevated levels of cAMP and defects in urinary concentrating ability by use of vasopressin receptor-2 antagonists such as tolvaptan; animal models paved the way for the use of this drug in clinical practice and in current trials of childhood AD-PKD [69]. Other approaches to treat NPHP have included CDK inihibitors, SHH agonists, and mTOR pathway inhibitors such as rapamycin, but many of these are in early stages of development [99]. Gene therapy holds some promise in treating cystic renal disease in JS due to CEP290 mutations; an antisense oligonucleotide (ASO) has been used to promote alternative splicing and exon skipping to rescue an intronic mutation in this gene in patient-derived renal epithelial cells and in a mouse model of JS, with reduction of cystic kidney disease burden [81]. Similarly, a comparable approach to rescuing the retinal phenotype of LCA caused by the common intronic splicing defect in the CEP290 gene using gene therapy approaches has demonstrated some success in a cell model and in mice carrying the mini-gene [29]. Finally, as other articles in this special issue point out, there are novel strategies emerging to treat these and other manifestations of ciliopathy conditions (“Retinal disease in ciliopathies: recent advances with a focus on stem cell-based therapies review” by Chen, H et al., “Novel Treatments for Polycystic Kidney Disease” by Patil, A et al., and “Using human urine-derived renal epithelial cells to model kidney disease in inherited ciliopathies” by Sayer, J and Molinari, E.).
Acknowledgments
The author thanks the many families who have participated in past and current studies of Joubert syndrome and related ciliary disorders, the Joubert Syndrome and Related Disorders Foundation for their support, and many professional collaborators, including Drs. Meral Gunay-Aygun, Ian Glass, and Daniel Doherty.
References
[1] | Akizu N. , Silhavy J.L. , Rosti R.O. , Scott E. , Fenstermaker A.G. , Schroth J. , Zaki M.S. , Sanchez H. , Gupta N. , Kabra M. , Kara M. , Ben-Omran T. , Rosti B. , Guemez-Gamboa A. , Spencer E. , Pan R. , Cai N. , Abdellateef M. , Gabriel S. , Halbritter J. , Hildebrandt F. , van Bokhoven H. , Gunel M. and Gleeson J.G. , Mutations in CSPP1 lead to classical Joubert syndrome, Am J Hum Genet 94: ((2014) ), 80–86. |
[2] | Alby C. , Piquand K. , Huber C. , Megarbane A. , Ichkou A. , Legendre M. , Pelluard F. , Encha-Ravazi F. , Abi-Tayeh G. , Bessieres B. , El Chehadeh-Djebbar S. , Laurent N. , Faivre L. , Sztriha L. , Zombor M. , Szabo H. , Failler M. , Garfa-Traore M. , Bole C. , Nitschke P. , Nizon M. , Elkhartoufi N. , Clerget-Darpoux F. , Munnich A. , Lyonnet S. , Vekemans M. , Saunier S. , Cormier-Daire V. , Attie-Bitach T. and Thomas S. , Mutations in KIAAcause lethal ciliopathies ranging from a hydrolethalus phenotype to short-rib polydactyly syndrome, Am J Hum Genet 97: ((2015) ), 311–318. |
[3] | Alkanderi S. , Molinari E. , Shaheen R. , Elmaghloob Y. , Stephen L.A. , Sammut V. , Ramsbottom S.A. , Srivastava S. , Cairns G. , Edwards N. , Rice S.J. , Ewida N. , Alhashem A. , White K. , Miles C.G. , Steel D.H. , Alkuraya F.S. , Ismail S. and Sayer J.A. , ARL3 mutations cause joubert syndrome by disrupting ciliary protein composition, Am J Hum Genet 103: ((2018) ), 612–620. |
[4] | Arts H. and Knoers N. , Cranioectodermal Dysplasia, 2013 Sep 12 [Updated 2018 Apr 12]. In: M.P. Adam, H.H. Ardinger, R.A. Pagon, et al., editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993–2019. |
[5] | Baala L. , Romano S. , Khaddour R. , Saunier S. , Smith U.M. , Audollent S. , Ozilou C. , Faivre L. , Laurent N. , Foliguet B. , Munnich A. , Lyonnet S. , Salomon R. , Encha-Razavi F. , Gubler M.C. , Boddaert N. , de Lonlay P. , Johnson C.A. , Vekemans M. , Antignac C. and Attie-Bitach T. , The meckel-gruber syndrome gene, MKS3, is mutated in Joubert syndrome, Am J Hum Genet 80: ((2007) ), 186–194. |
[6] | Bachmann-Gagescu R. , Dempsey J.C. , Phelps I.G. , O’Roak B.J. , Knutzen D.M. , Rue T.C. , Ishak G.E. , Isabella C.R. , Gorden N. , Adkins J. , Boyle E.A. , de Lacy N. , O’Day D. , Alswaid A. , Ramadevi A.R. , Lingappa L. , Lourenco C. , Martorell L. , Garcia-Cazorla A. , Ozyurek H. , Haliloglu G. , Tuysuz B. , Topcu M. , University of Washington Center for Mendelian G. , Chance P. , Parisi M.A. , Glass I.A. , Shendure J. , Doherty D. , Joubert syndrome: A model for untangling recessive disorders with extreme genetic heterogeneity, J Med Genet 52: ((2015) ), 514–522. |
[7] | Bachmann-Gagescu R. , Dona M. , Hetterschijt L. , Tonnaer E. , Peters T. , de Vrieze E. Mans D.A. , van Beersum S.E. , Phelps I.G. , Arts H.H. , Keunen J.E. , Ueffing M. , Roepman R. , Boldt K. , Doherty D. , Moens C.B. , Neuhauss S.C. , Kremer H. and van Wijk E. , The ciliopathy protein CC2D2A associates with NINL and functions in RAB8-MICAL3-regulated vesicle trafficking, PLoS Genet 11: ((2015) ), e1005575. |
[8] | Bachmann-Gagescu R. , Ishak G.E. , Dempsey J.C. , Adkins J. , O’Day D. , Phelps I.G. , Gunay-Aygun M. , Kline A.D. , Szczaluba K. , Martorell L. , Alswaid A. , Alrasheed S. , Pai S. , Izatt L. , Ronan A. , Parisi M.A. , Mefford H. , Glass I. , Doherty D. , Genotype-phenotype correlation in CC2D2A-related Joubert syndrome reveals an association with ventriculomegaly and seizures, J Med Genet 49: ((2012) ), 126–137. |
[9] | Bachmann-Gagescu R. , Phelps I.G. , Dempsey J.C. , Sharma V.A. , Ishak G.E. , Boyle E.A. , Wilson M. , Marques Lourenco C. Arslan M. , University of Washington Center for Mendelian G. , Shendure J. and Doherty D. , KIAAis Mutated in Joubert Syndrome, Hum Mutat 36: ((2015) ), 831–835. |
[10] | Beck B.B. , Phillips J.B. , Bartram M.P. , Wegner J. , Thoenes M. , Pannes A. , Sampson J. , Heller R. , Gobel H. , Koerber F. , Neugebauer A. , Hedergott A. , Nurnberg G. , Nurnberg P. , Thiele H. , Altmuller J. , Toliat M.R. , Staubach S. , Boycott K.M. , Valente E.M. , Janecke A.R. , Eisenberger T. , Bergmann C. , Tebbe L. , Wang Y. , Wu Y. , Fry A.M. , Westerfield M. , Wolfrum U. and Bolz H.J. , Mutation of POC1B in a severe syndromic retinal ciliopathy, Hum Mutat 35: ((2014) ), 1153–1162. |
[11] | Bielas S.L. , Silhavy J.L. , Brancati F. , Kisseleva M.V. , Al-Gazali L. , Sztriha L. , Bayoumi R.A. , Zaki M.S. , Abdel-Aleem A. , Rosti R.O. , Kayserili H. , Swistun D. , Scott L.C. , Bertini E. , Boltshauser E. , Fazzi E. , Travaglini L. , Field S.J. , Gayral S. , Jacoby M. , Schurmans S. , Dallapiccola B. , Majerus P.W. , Valente E.M. and Gleeson J.G. , Mutations in INPP5E, encoding inositol polyphosphate-5-phosphatase E, link phosphatidyl inositol signaling to the ciliopathies, Nat Genet 41: ((2009) ), 1032–1036. |
[12] | Bonnard C. , Shboul M. , Tonekaboni S.H. , Ng A.Y.J. , Tohari S. , Ghosh K. , Lai A. , Lim J.Y. , Tan E.C. , Devisme L. , Stichelbout M. , Alkindi A. , Banu N. , Yuksel Z. , Ghoumid J. , Elkhartoufi N. , Boutaud L. , Micalizzi A. , Brett M.S. , Venkatesh B. , Valente E.M. , Attie-Bitach T. , Reversade B. and Kariminejad A. , Novel mutations in the ciliopathy-associated gene CPLANE1 (C5orf42) cause OFD syndrome type VI rather than Joubert syndrome, Eur J Med Genet 61: ((2018) ), 585–595. |
[13] | Brancati F. , Barrano G. , Silhavy J.L. , Marsh S.E. , Travaglini L. , Bielas S.L. , Amorini M. , Zablocka D. , Kayserili H. , Al-Gazali L. , Bertini E. , Boltshauser E. , D’Hooghe M. , Fazzi E. , Fenerci E.Y. , Hennekam R.C. , Kiss A. , Lees M.M. , Marco E. , Phadke S.R. , Rigoli L. , Romano S. , Salpietro C.D. , Sherr E.H. , Signorini S. , Stromme P. , Stuart B. , Sztriha L. , Viskochil D.H. , Yuksel A. , Dallapiccola B. , International J.S.G. , Valente E.M. and Gleeson J.G. , CEP290 mutations are frequently identified in the oculo-renal form of Joubert syndrome-related disorders, Am J Hum Genet 81: ((2007) ), 104–113. |
[14] | Brancati F. , Dallapiccola B. and Valente E.M. , Joubert Syndrome and related disorders, Orphanet J Rare Dis 5: ((2010) ), 20. |
[15] | Brancati F. , Iannicelli M. , Travaglini L. , Mazzotta A. , Bertini E. , Boltshauser E. , D’Arrigo S. , Emma F. , Fazzi E. , Gallizzi R. , Gentile M. , Loncarevic D. , Mejaski-Bosnjak V. , Pantaleoni C. , Rigoli L. , Salpietro C.D. , Signorini S. , Stringini G.R. , Verloes A. , Zabloka D. , Dallapiccola B. , Gleeson J.G. , Valente E.M. and International J.S.G. , MKS3/TMEM67 mutations are a major cause of COACH Syndrome, a Joubert Syndrome related disorder with liver involvement, Hum Mutat 30: ((2009) ), E432–E442. |
[16] | Brooks B.P. , Zein W.M. , Thompson A.H. , Mokhtarzadeh M. , Doherty D.A. , Parisi M. , Glass I.A. , Malicdan M.C. , Vilboux T. , Vemulapalli M. , Mullikin J.C. , Gahl W.A. and Gunay-Aygun M. , Joubert syndrome: Ophthalmological findings in correlation with genotype and hepatorenal disease in 99 patients prospectively evaluated at a single center, Ophthalmology 125: ((2018) ), 1937–1952. |
[17] | Cantagrel V. , Silhavy J.L. , Bielas S.L. , Swistun D. , Marsh S.E. , Bertrand J.Y. , Audollent S. , Attie-Bitach T. , Holden K.R. , Dobyns W.B. , Traver D. , Al-Gazali L. , Ali B.R. , Lindner T.H. , Caspary T. , Otto E.A. , Hildebrandt F. , Glass I.A. , Logan C.V. , Johnson C.A. , Bennett C. , Brancati F. , International Joubert Syndrome Related Disorders Study G. , Valente E.M. , Woods C.G. , Gleeson J.G. , Mutations in the cilia gene ARL13B lead to the classical form of Joubert syndrome, Am J Hum Genet 83: ((2008) ), 170–179. |
[18] | Chaki M. , Airik R. , Ghosh A.K. , Giles R.H. , Chen R. , Slaats G.G. , Wang H. , Hurd T.W. , Zhou W. , Cluckey A. , Gee H.Y. , Ramaswami G. , Hong C.J. , Hamilton B.A. , Cervenka I. , Ganji R.S. , Bryja V. , Arts H.H. , van Reeuwijk J. Oud M.M. , Letteboer S.J. , Roepman R. , Husson H. , Ibraghimov-Beskrovnaya O. , Yasunaga T. , Walz G. , Eley L. , Sayer J.A. , Schermer B. , Liebau M.C. , Benzing T. , Le Corre S. , Drummond I. , Janssen S. , Allen S.J. , Natarajan S. , O’Toole J.F. , Attanasio M. , Saunier S. , Antignac C. , Koenekoop R.K. , Ren H. , Lopez I. , Nayir A. , Stoetzel C. , Dollfus H. , Massoudi R. , Gleeson J.G. , Andreoli S.P. , Doherty D.G. , Lindstrad A. , Golzio C. , Katsanis N. , Pape L. , Abboud E.B. , Al-Rajhi A.A. , Lewis R.A. , Omran R.A.H. , Lee E.Y. , Wang S. , Sekiguchi J.M. , Saunders R. , Johnson C.A. , Garner E. , Vanselow K. , Andersen J.S. , Shlomai J. , Nurnberg G. , Levy S. , Smogorzewska A. , Otto E.A. and Hildebrandt F. , Exome capture reveals ZNF423 and CEP164 mutations, linking renal ciliopathies to DNA damage response signaling, Cell 150: ((2012) ), 533–548. |
[19] | Chih B. , Liu P. , Chinn Y. , Chalouni C. , Komuves L.G. , Hass P.E. , Sandoval W. , Peterson A.S. , A ciliopathy complex at the transition zone protects the cilia as a privileged membrane domain, Nat Cell Biol 14: ((2011) ), 61–72. |
[20] | Coene K.L. , Roepman R. , Doherty D. , Afroze B. , Kroes H.Y. , Letteboer S.J. , Ngu L.H. , Budny B. , van Wijk E. , Gorden N.T. , Azhimi M. , Thauvin-Robinet C. , Veltman J.A. , Boink M. , Kleefstra T. , Cremers F.P. , van Bokhoven H. and de Brouwer A.P. , OFD1 is mutated in X-linked Joubert syndrome and interacts with LCA5-encoded lebercilin, Am J Hum Genet 85: ((2009) ), 465–481. |
[21] | Dafinger C. , Liebau M.C. , Elsayed S.M. , Hellenbroich Y. , Boltshauser E. , Korenke G.C. , Fabretti F. , Janecke A.R. , Ebermann I. , Nurnberg G. , Nurnberg P. , Zentgraf H. , Koerber F. , Addicks K. , Elsobky E. , Benzing T. , Schermer B. , Bolz H.J. , Mutations in KIF7 link Joubert syndrome with Sonic Hedgehog signaling and microtubule dynamics, J Clin Invest 121: ((2011) ), 2662–2667. |
[22] | De Mori R. , Romani M. , D’Arrigo S. , Zaki M.S. , Lorefice E. , Tardivo S. , Biagini T. , Stanley V. , Musaev D. , Fluss J. , Micalizzi A. , Nuovo S. , Illi B. , Chiapparini L. , Di Marcotullio L. , Issa M.Y. , Anello D. , Casella A. , Ginevrino M. , Leggins A.S. , Roosing S. , Alfonsi R. , Rosati J. , Schot R. , Mancini G.M.S. , Bertini E. , Dobyns W.B. , Mazza T. , Gleeson J.G. and Valente E.M. , Hypomorphic Recessive Variants in SUFU Impair the Sonic Hedgehog Pathway and Cause Joubert Syndrome with Cranio-facial and Skeletal Defects, Am J Hum Genet 101: ((2017) ), 552–563. |
[23] | Delous M. , Baala L. , Salomon R. , Laclef C. , Vierkotten J. , Tory K. , Golzio C. , Lacoste T. , Besse L. , Ozilou C. , Moutkine I. , Hellman N.E. , Anselme I. , Silbermann F. , Vesque C. , Gerhardt C. , Rattenberry E. , Wolf M.T. , Gubler M.C. , Martinovic J. , Encha-Razavi F. , Boddaert N. , Gonzales M. , Macher M.A. , Nivet H. , Champion G. , Bertheleme J.P. , Niaudet P. , McDonald F. , Hildebrandt F. , Johnson C.A. , Vekemans M. , Antignac C. , Ruther U. , Schneider-Maunoury S. , Attie-Bitach T. and Saunier S. , The ciliary gene RPGRIP1L is mutated in cerebello-oculo-renal syndrome (Joubert syndrome type B) and Meckel syndrome, Nat Genet 39: ((2007) ), 875–881. |
[24] | Dempsey J.C. , Phelps I.G. , Bachmann-Gagescu R. , Glass I.A. , Tully H.M. and Doherty D. , Mortality in Joubert syndrome, Am J Med Genet A 173: ((2017) ), 1237–1242. |
[25] | Dixon-Salazar T. , Silhavy J.L. , Marsh S.E. , Louie C.M. , Scott L.C. , Gururaj A. , Al-Gazali L. , Al-Tawari A.A. , Kayserili H. , Sztriha L. and Gleeson J.G. , Mutations in the AHI1 gene, encoding jouberin, cause Joubert syndrome with cortical polymicrogyria, Am J Hum Genet 75: ((2004) ), 979–987. |
[26] | Doherty D. , Joubert syndrome: Insights into brain development, cilium biology, and complex disease, Semin Pediatr Neurol 16: ((2009) ), 143–154. |
[27] | Doherty D. , Glass I.A. , Siebert J.R. , Strouse P.J. , Parisi M.A. , Shaw D.W. , Chance P.F. , Barr M. Jr and Nyberg D. , Prenatal diagnosis in pregnancies at risk for Joubert syndrome by ultrasound and MRI, Prenat Diagn 25: ((2005) ), 442–447. |
[28] | Doherty D. , Parisi M.A. , Finn L.S. , Gunay-Aygun M. , Al-Mateen M. , Bates D. , Clericuzio C. , Demir H. , Dorschner M. , van Essen A.J. , Gahl W.A. , Gentile M. , Gorden N.T. , Hikida A. , Knutzen D. , Ozyurek H. , Phelps I. , Rosenthal P. , Verloes A. , Weigand H. , Chance P.F. , Dobyns W.B. and Glass I.A. , Mutations in 3 genes (MKS3, CC2D2A and RPGRIP1L) cause COACH syndrome (Joubert syndrome with congenital hepatic fibrosis), J Med Genet 47: ((2010) ), 8–21. |
[29] | Dooley S.J. , McDougald D.S. , Fisher K.J. , Bennicelli J.L. , Mitchell L.G. and Bennett J. , Spliceosome-mediated Pre-mRNA trans-splicing can repair CEP290 mRNA, Mol Ther Nucleic Acids 12: ((2018) ), 294–308. |
[30] | Edvardson S. , Shaag A. , Zenvirt S. , Erlich Y. , Hannon G.J. , Shanske A.L. , Gomori J.M. , Ekstein J. and Elpeleg O. , Joubert syndrome 2 (JBTS2) in Ashkenazi Jews is associated with a TMEM216 mutation, Am J Hum Genet 86: ((2010) ), 93–97. |
[31] | Enokizono M. , Aida N. , Niwa T. , Osaka H. , Naruto T. , Kurosawa K. , Ohba C. , Suzuki T. , Saitsu H. , Goto T. and Matsumoto N. , Neuroimaging findings in Joubert syndrome with C5orf42 gene mutations: A milder form of molar tooth sign and vermian hypoplasia, J Neurol Sci 376: ((2017) ), 7–12. |
[32] | Field M. , Scheffer I.E. , Gill D. , Wilson M. , Christie L. , Shaw M. , Gardner A. , Glubb G. , Hobson L. , Corbett M. , Friend K. , Willis-Owen S. and Gecz J. , Expanding the molecular basis and phenotypic spectrum of X-linked Joubert syndrome associated with OFD1 mutations, Eur J Hum Genet 20: ((2012) ), 806–809. |
[33] | Fleming L.R. , Doherty D.A. , Parisi M.A. , Glass I.A. , Bryant J. , Fischer R. , Turkbey B. , Choyke P. , Daryanani K. , Vemulapalli M. , Mullikin J.C. , Malicdan M.C. , Vilboux T. , Sayer J.A. , Gahl W.A. and Gunay-Aygun M. , Prospective evaluation of kidney disease in joubert syndrome, Clin J Am Soc Nephrol 12: ((2017) ), 1962–1973. |
[34] | Forsythe E. and Beales P.L. ((2015) ) Bardet-Biedl Syndrome, 2003 Jul 14 [Updated 2015 Apr 23]., In: M.Adam, H.H. Ardinger, R.A. Pagon, et al., editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993–2019. |
[35] | Garcia-Gonzalo F.R. , Corbit K.C. , Sirerol-Piquer M.S. , Ramaswami G. , Otto E.A. , Noriega T.R. , Seol A.D. , Robinson J.F. , Bennett C.L. , Josifova D.J. , Garcia-Verdugo J.M. , Katsanis N. , Hildebrandt F. and Reiter J.F. , A transition zone complex regulates mammalian ciliogenesis and ciliary membrane composition, Nat Genet 43: ((2011) ), 776–784. |
[36] | Garcia-Gonzalo F.R. and Reiter J.F. , Open sesame: How transition fibers and the transition zone control ciliary composition, Cold Spring Harb Perspect Biol 9: ((2017) ). |
[37] | Gleeson J.G. , Keeler L.C. , Parisi M.A. , Marsh S.E. , Chance P.F. , Glass I.A. , Graham J.M. Jr., , Maria B.L. , Barkovich A.J. and Dobyns W.B. , Molar tooth sign of the midbrain-hindbrain junction: Occurrence in multiple distinct syndromes, Am J Med Genet A 125A: ((2004) ), 125–134; discussion 117. |
[38] | Goncalves J. and Pelletier L. , The ciliary transition zone: Finding the pieces and assembling the gate, Mol Cells 40: ((2017) ), 243–253. |
[39] | Gorden N.T. , Arts H.H. , Parisi M.A. , Coene K.L. , Letteboer S.J. , van Beersum S.E. , Mans D.A. , Hikida A. , Eckert M. , Knutzen D. , Alswaid A.F. , Ozyurek H. , Dibooglu S. , Otto E.A. , Liu Y. , Davis E.E. , Hutter C.M. , Bammler T.K. , Farin F.M. , Dorschner M. , Topcu M. , Zackai E.H. , Rosenthal P. , Owens K.N. , Katsanis N. , Vincent J.B. , Hildebrandt F. , Rubel E.W. , Raible D.W. , Knoers N.V. , Chance P.F. , Roepman R. , Moens C.B. , Glass I.A. and Doherty D. , CC2D2A is mutated in Joubert syndrome and interacts with the ciliopathy-associated basal body protein CEP290, Am J Hum Genet 83: ((2008) ), 559–571. |
[40] | Gunay-Aygun M. , Liver and kidney disease in ciliopathies, Am J Med Genet C Semin Med Genet 151C: ((2009) ), 296–306. |
[41] | Gunay-Aygun M. , Parisi M.A. , Doherty D. , Tuchman M. , Tsilou E. , Kleiner D.E. , Huizing M. , Turkbey B. , Choyke P. , Guay-Woodford L. , Heller T. , Szymanska K. , Johnson C.A. , Glass I. and Gahl W.A. , MKS3-related ciliopathy with features of autosomal recessive polycystic kidney disease, nephronophthisis, and Joubert Syndrome, J Pediatr 155: ((2009) ), 386–392 e381. |
[42] | Halbritter J. , Bizet A.A. , Schmidts M. , Porath J.D. , Braun D.A. , Gee H.Y. , McInerney-Leo A.M. , Krug P. , Filhol E. , Davis E.E. , Airik R. , Czarnecki P.G. , Lehman A.M. , Trnka P. , Nitschke P. , Bole-Feysot C. , Schueler M. , Knebelmann B. , Burtey S. , Szabo A.J. , Tory K. , Leo P.J. , Gardiner B. , McKenzie F.A. , Zankl A. , Brown M.A. , Hartley J.L. , Maher E.R. , Li C. , Leroux M.R. , Scambler P.J. , Zhan S.H. , Jones S.J. , Kayserili H. , Tuysuz B. , Moorani K.N. , Constantinescu A. , Krantz I.D. , Kaplan B.S. , Shah J.V. , Consortium U.K. , Hurd T.W. , Doherty D. , Katsanis N. , Duncan E.L. , Otto E.A. , Beales P.L. , Mitchison H.M. , Saunier S. and Hildebrandt F. , Defects in the IFT-B component IFT172 cause Jeune and Mainzer-Saldino syndromes in humans, Am J Hum Genet 93: ((2013) ), 915–925. |
[43] | Hartill V. , Szymanska K. , Sharif S.M. , Wheway G. and Johnson C.A. , Meckel-gruber syndrome: An update on diagnosis, clinical management, and research advances, Front Pediatr 5: ((2017) ), 244. |
[44] | Hildebrandt F. , Attanasio M. and Otto E. , Nephronophthisis: Disease mechanisms of a ciliopathy, J Am Soc Nephrol 20: ((2009) ), 23–35. |
[45] | Hildebrandt F. , Strahm B. , Nothwang H.G. , Gretz N. , Schnieders B. , Singh-Sawhney I. , Kutt R. , Vollmer M. and Brandis M. , Molecular genetic identification of families with juvenile nephronophthisis type 1: Rate of progression to renal failure. APN Study Group. Arbeitsgemeinschaft fur Padiatrische Nephrologie, Kidney Int 51: ((1997) ), 261–269. |
[46] | Hoefele J. , Wolf M.T. , O’Toole J.F. , Otto E.A. , Schultheiss U. , Deschenes G. , Attanasio M. , Utsch B. , Antignac C. and Hildebrandt F. , Evidence of oligogenic inheritance in nephronophthisis, J Am Soc Nephrol 18: ((2007) ), 2789–2795. |
[47] | Huang L. , Szymanska K. , Jensen V.L. , Janecke A.R. , Innes A.M. , Davis E.E. , Frosk P. , Li C. , Willer J.R. , Chodirker B.N. , Greenberg C.R. , McLeod D.R. , Bernier F.P. , Chudley A.E. , Muller T. , Shboul M. , Logan C.V. , Loucks C.M. , Beaulieu C.L. , Bowie R.V. , Bell S.M. , Adkins J. , Zuniga F.I. , Ross K.D. , Wang J. , Ban M.R. , Becker C. , Nurnberg P. , Douglas S. , Craft C.M. , Akimenko M.A. , Hegele R.A. , Ober C. , Utermann G. , Bolz H.J. , Bulman D.E. , Katsanis N. , Blacque O.E. , Doherty D. , Parboosingh J.S. , Leroux M.R. , Johnson C.A. and Boycott K.M. , TMEM237 is mutated in individuals with a Joubert syndrome related disorder and expands the role of the TMEM family at the ciliary transition zone, Am J Hum Genet 89: ((2011) ), 713–730. |
[48] | Jacoby M. , Cox J.J. , Gayral S. , Hampshire D.J. , Ayub M. , Blockmans M. , Pernot E. , Kisseleva M.V. , Compere P. , Schiffmann S.N. , Gergely F. , Riley J.H. , Perez-Morga D. , Woods C.G. and Schurmans S. , INPP5E mutations cause primary cilium signaling defects, ciliary instability and ciliopathies in human and mouse, Nat Genet 41: ((2009) ), 1027–1031. |
[49] | Joubert M. , Eisenring J.J. , Robb J.P. and Andermann F. , Familial agenesis of the cerebellar vermis. A syndrome of episodic hyperpnea, abnormal eye movements, ataxia, and retardation, Neurology 19: ((1969) ), 813–825. |
[50] | Katsanis N. , Ansley S.J. , Badano J.L. , Eichers E.R. , Lewis R.A. , Hoskins B.E. , Scambler P.J. , Davidson W.S. , Beales P.L. and Lupski J.R. , Triallelic inheritance in Bardet-Biedl syndrome, a Mendelian recessive disorder, Science 293: ((2001) ), 2256–2259. |
[51] | Khaddour R. , Smith U. , Baala L. , Martinovic J. , Clavering D. , Shaffiq R. , Ozilou C. , Cullinane A. , Kyttala M. , Shalev S. , Audollent S. , d’Humieres C. , Kadhom N. , Esculpavit C. , Viot G. , Boone C. , Oien C. , Encha-Razavi F. , Batman P.A. , Bennett C.P. , Woods C.G. , Roume J. , Lyonnet S. , Genin E. , Le Merrer M. , Munnich A. , Gubler M.C. , Cox P. , Macdonald F. , Vekemans M. , Johnson C.A. , Attie-Bitach T. and Soffoet , Spectrum of MKS1 and MKS3 mutations in Meckel syndrome: A genotype-phenotype correlation, Mutation in brief #960. Online, Hum Mutat 28: , ((2007) ), 523–524. |
[52] | Khanna H. , Davis E.E. , Murga-Zamalloa C.A. , Estrada-Cuzcano A. , Lopez I. , den Hollander A.I. , Zonneveld M.N. , Othman M.I. , Waseem N. , Chakarova C.F. , Maubaret C. , Diaz-Font A. , MacDonald I. , Muzny D.M. , Wheeler D.A. , Morgan M. , Lewis L.R. , Logan C.V. , Tan P.L. , Beer M.A. , Inglehearn C.F. , Lewis R.A. , Jacobson S.G. , Bergmann C. , Beales P.L. , Attie-Bitach T. , Johnson C.A. , Otto E.A. , Bhattacharya S.S. , Hildebrandt F. , Gibbs R.A. , Koenekoop R.K. , Swaroop A. and Katsanis N. , A common allele in RPGRIP1L is a modifier of retinal degeneration in ciliopathies, Nat Genet 41: ((2009) ), 739–745. |
[53] | Kroes H.Y. , Monroe G.R. , van der Zwaag B. , Duran K.J. , de Kovel C.G. , van Roosmalen M.J. , Harakalova M. , Nijman I.J. , Kloosterman W.P. , Giles R.H. , Knoers N.V. and van Haaften G. , Joubert syndrome: Genotyping a Northern European patient cohort, Eur J Hum Genet 24: ((2016) ), 214–220. |
[54] | Kumaran N. , Pennesi M.E. , Yang P. , Trzupek K.M. , Schlechter C. , Moore A.T. , Weleber R.G. and Michaelides M. ((2018) ) Leber Congenital Amaurosis / Early-Onset Severe Retinal Dystrophy Overview, 2018 Oct 4. In: M.P. Adam, H.H. Ardinger, R.A. Pagon, et al., editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle;. 1993–2019. |
[55] | Lambacher N.J. , Bruel A.L. , van Dam T.J. , Szymanska K. , Slaats G.G. , Kuhns S. , McManus G.J. , Kennedy J.E. , Gaff K. , Wu K.M. , van der Lee R. , Burglen L. , Doummar D. , Riviere J.B. , Faivre L. , Attie-Bitach T. , Saunier S. , Curd A. , Peckham M. , Giles R.H. , Johnson C.A. , Huynen M.A. , Thauvin-Robinet C. and Blacque O.E. , TMEM107 recruits ciliopathy proteins to subdomains of the ciliary transition zone and causes Joubert syndrome, Nat Cell Biol 18: ((2016) ), 122–131. |
[56] | Lee J.E. , Silhavy J.L. , Zaki M.S. , Schroth J. , Bielas S.L. , Marsh S.E. , Olvera J. , Brancati F. , Iannicelli M. , Ikegami K. , Schlossman A.M. , Merriman B. , Attie-Bitach T. , Logan C.V. , Glass I.A. , Cluckey A. , Louie C.M. , Lee J.H. , Raynes H.R. , Rapin I. , Castroviejo I.P. , Setou M. , Barbot C. , Boltshauser E. , Nelson S.F. , Hildebrandt F. , Johnson C.A. , Doherty D.A. , Valente E.M. and Gleeson J.G. , CEP41 is mutated in Joubert syndrome and is required for tubulin glutamylation at the cilium, Nat Genet 44: ((2012) ), 193–199. |
[57] | Lee J.H. , Silhavy J.L. , Lee J.E. , Al-Gazali L. , Thomas S. , Davis E.E. , Bielas S.L. , Hill K.J. , Iannicelli M. , Brancati F. , Gabriel S.B. , Russ C. , Logan C.V. , Sharif S.M. , Bennett C.P. , Abe M. , Hildebrandt F. , Diplas B.H. , Attie-Bitach T. , Katsanis N. , Rajab A. , Koul R. , Sztriha L. , Waters E.R. , Ferro-Novick S. , Woods C.G. , Johnson C.A. , Valente E.M. , Zaki M.S. and Gleeson J.G. , Evolutionarily assembled cis-regulatory module at a human ciliopathy locus, Science 335: ((2012) ), 966–969. |
[58] | Lehman A.M. , Eydoux P. , Doherty D. , Glass I.A. , Chitayat D. , Chung B.Y. , Langlois S. , Yong S.L. , Lowry R.B. , Hildebrandt F. and Trnka P. , Co-occurrence of Joubert syndrome and Jeune asphyxiating thoracic dystrophy, Am J Med Genet A 152A: ((2010) ), 1411–1419. |
[59] | Lindstrand A. , Frangakis S. , Carvalho C.M. , Richardson E.B. , McFadden K.A. , Willer J.R. , Pehlivan D. , Liu P. , Pediaditakis I.L. , Sabo A. , Lewis R.A. , Banin E. , Lupski J.R. , Davis E.E. and Katsanis N. , Copy-number variation contributes to the mutational load of bardet-biedl syndrome, Am J Hum Genet 99: ((2016) ), 318–336. |
[60] | Lopez E. , Thauvin-Robinet C. , Reversade B. , Khartoufi N.E. , Devisme L. , Holder M. , Ansart-Franquet H. , Avila M. , Lacombe D. , Kleinfinger P. , Kaori I. , Takanashi J. , Le Merrer M. , Martinovic J. , Noel C. , Shboul M. , Ho L. , Guven Y. , Razavi F. , Burglen L. , Gigot N. , Darmency-Stamboul V. , Thevenon J. , Aral B. , Kayserili H. , Huet F. , Lyonnet S. , Le Caignec C. , Franco B. , Riviere J.B. , Faivre L. , Attie-Bitach T. , C5orf42 is the major gene responsible for OFD syndrome type VI, Hum Genet 133: ((2014) ), 367–377. |
[61] | Maglic D. , Stephen J. , Malicdan M.C. , Guo J. , Fischer R. , Konzman D. , Program N.C.S. , Mullikin J.C. , Gahl W.A. , Vilboux T. and Gunay-Aygun M. , TMEM231 gene conversion associated with joubert and meckel-gruber syndromes in the same family, Hum Mutat 37: ((2016) ), 1144–1148. |
[62] | Malicdan M.C. , Vilboux T. , Stephen J. , Maglic D. , Mian L. , Konzman D. , Guo J. , Yildirimli D. , Bryant J. , Fischer R. , Zein W.M. , Snow J. , Vemulapalli M. , Mullikin J.C. , Toro C. , Solomon B.D. , Niederhuber J.E. , Program N.C.S. , Gahl W.A. and Gunay-Aygun M. , Mutations in human homologue of chicken talpid3 gene (KIAAcause a hybrid ciliopathy with overlapping features of Jeune and Joubert syndromes, J Med Genet 52: ((2015) ), 830–839. |
[63] | Maria B.L. , Boltshauser E. , Palmer S.C. and Tran T.X. , Clinical features and revised diagnostic criteria in Joubert syndrome, J Child Neurol 14: ((1999) ), 583–590; discussion 590–581. |
[64] | Maria B.L. , Hoang K.B. , Tusa R.J. , Mancuso A.A. , Hamed L.M. , Quisling R.G. , Hove M.T. , Fennell E.B. , Booth-Jones M. , Ringdahl D.M. , Yachnis A.T. , Creel G. and Frerking B. , “Joubert syndrome” revisited: Key ocular motor signs with magnetic resonance imaging correlation, J Child Neurol 12: ((1997) ), 423–430. |
[65] | Maria B.L. , Quisling R.G. , Rosainz L.C. , Yachnis A.T. , Gitten J. , Dede D. and Fennell E. , Molar tooth sign in Joubert syndrome: Clinical, radiologic, and pathologic significance, J Child Neurol 14: ((1999) ), 368–376. |
[66] | Mitchison H.M. and Valente E.M. , Motile and non-motile cilia in human pathology: From function to phenotypes, J Pathol 241: ((2017) ), 294–309. |
[67] | Moore S.J. , Green J.S. , Fan Y. , Bhogal A.K. , Dicks E. , Fernandez B.A. , Stefanelli M. , Murphy C. , Cramer B.C. , Dean J.C. , Beales P.L. , Katsanis N. , Bassett A.S. , Davidson W.S. and Parfrey P.S. , Clinical and genetic epidemiology of Bardet-Biedl syndrome in Newfoundland: A 22-year prospective, population-based, cohort study, Am J Med Genet A 132A: ((2005) ), 352–360. |
[68] | Mougou-Zerelli S. , Thomas S. , Szenker E. , Audollent S. , Elkhartoufi N. , Babarit C. , Romano S. , Salomon R. , Amiel J. , Esculpavit C. , Gonzales M. , Escudier E. , Leheup B. , Loget P. , Odent S. , Roume J. , Gerard M. , Delezoide A.L. , Khung S. , Patrier S. , Cordier M.P. , Bouvier R. , Martinovic J. , Gubler M.C. , Boddaert N. , Munnich A. , Encha-Razavi F. , Valente E.M. , Saad A. , Saunier S. , Vekemans M. and Attie-Bitach T. , CC2D2A mutations in Meckel and Joubert syndromes indicate a genotype-phenotype correlation, Hum Mutat 30: ((2009) ), 1574–1582. |
[69] | Muller R.U. , Haas C.S. and Sayer J.A. , Practical approaches to the management of autosomal dominant polycystic kidney disease patients in the era of tolvaptan, Clin Kidney J 11: ((2018) ), 62–69. |
[70] | Parisi M. and Glass I. ((2017) ) Joubert Syndrome, 2003 Jul 9 [Updated 2017 Jun 29]. In: M.Adam, H.H. Ardinger, R.A. Pagon, et al., editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993–2019. |
[71] | Parisi M.A. , Clinical and molecular features of Joubert syndrome and related disorders, Am J Med Genet C Semin Med Genet 151C: ((2009) ), 326–340. |
[72] | Parisi M.A. , Bennett C.L. , Eckert M.L. , Dobyns W.B. , Gleeson J.G. , Shaw D.W. , McDonald R. , Eddy A. , Chance P.F. and Glass I.A. , The NPHP1 gene deletion associated with juvenile nephronophthisis is present in a subset of individuals with Joubert syndrome, Am J Hum Genet 75: ((2004) ), 82–91. |
[73] | Parisi M.A. , Doherty D. , Chance P.F. and Glass I.A. , Joubert syndrome (and related disorders) (OMIM 00), Eur J Hum Genet 15: ((2007) ), 511–521. |
[74] | Parisi M.A. , Doherty D. , Eckert M.L. , Shaw D.W. , Ozyurek H. , Aysun S. , Giray O. , Al Swaid A. , Al Shahwan S. , Dohayan N. , Bakhsh E. , Indridason O.S. , Dobyns W.B. , Bennett C.L. , Chance P.F. and Glass I.A. , AHI1 mutations cause both retinal dystrophy and renal cystic disease in Joubert syndrome, J Med Genet 43: ((2006) ), 334–339. |
[75] | Phelps I.G. , Dempsey J.C. , Grout M.E. , Isabella C.R. , Tully H.M. , Doherty D. and Bachmann-Gagescu R. , Interpreting the clinical significance of combined variants in multiple recessive disease genes: Systematic investigation of Joubert syndrome yields little support for oligogenicity, Genet Med 20: ((2018) ), 223–233. |
[76] | Poretti A. , Dietrich Alber F. , Brancati F. , Dallapiccola B. , Valente E.M. and Boltshauser E. , Normal cognitive functions in joubert syndrome, Neuropediatrics 40: ((2009) ), 287–290. |
[77] | Poretti A. , Snow J. , Summers A.C. , Tekes A. , Huisman T.A. , Aygun N. , Carson K.A. , Doherty D. , Parisi M.A. , Toro C. , Yildirimli D. , Vemulapalli M. , Mullikin J.C. , Program N.C.S. , Cullinane A.R. , Vilboux T. , Gahl W.A. and Gunay-Aygun M. , Joubert syndrome: Neuroimaging findings in 110 patients in correlation with cognitive function and genetic cause, J Med Genet ((2017) ). |
[78] | Pretorius P.R. , Baye L.M. , Nishimura D.Y. , Searby C.C. , Bugge K. , Yang B. , Mullins R.F. , Stone E.M. , Sheffield V.C. and Slusarski D.C. , Identification and functional analysis of the vision-specific BBS3 (ARL6) long isoform, PLoS Genet 6: ((2010) ), e1000884. |
[79] | Putoux A. , Thomas S. , Coene K.L. , Davis E.E. , Alanay Y. , Ogur G. , Uz E. , Buzas D. , Gomes C. , Patrier S. , Bennett C.L. , Elkhartoufi N. , Frison M.H. , Rigonnot L. , Joye N. , Pruvost S. , Utine G.E. , Boduroglu K. , Nitschke P. , Fertitta L. , Thauvin-Robinet C. , Munnich A. , Cormier-Daire V. , Hennekam R. , Colin E. , Akarsu N.A. , Bole-Feysot C. , Cagnard N. , Schmitt A. , Goudin N. , Lyonnet S. , Encha-Razavi F. , Siffroi J.P. , Winey M. , Katsanis N. , Gonzales M. , Vekemans M. , Beales P.L. and Attie-Bitach T. , KIF7 mutations cause fetal hydrolethalus and acrocallosal syndromes, Nat Genet 43: ((2011) ), 601–606. |
[80] | Quarello E. , Molho M. , Garel C. , Couture A. , Legac M.P. , Moutard M.L. , Bault J.P. , Fallet-Bianco C. and Guibaud L. , Prenatal abnormal features of the fourth ventricle in Joubert syndrome and related disorders, Ultrasound Obstet Gynecol 43: ((2014) ), 227–232. |
[81] | Ramsbottom S.A. , Molinari E. , Srivastava S. , Silberman F. , Henry C. , Alkanderi S. , Devlin L.A. , White K. , Steel D.H. , Saunier S. , Miles C.G. and Sayer J.A. , Targeted exon skipping of a CEP290 mutation rescues Joubert syndrome phenotypesand in a murine model, Proc Natl Acad Sci U S A 115: ((2018) ), 12489–12494. |
[82] | Reiter J.F. and Leroux M.R. , Genes and molecular pathways underpinning ciliopathies, Nat Rev Mol Cell Biol 18: ((2017) ), 533–547. |
[83] | Romani M. , Micalizzi A. , Kraoua I. , Dotti M.T. , Cavallin M. , Sztriha L. , Ruta R. , Mancini F. , Mazza T. , Castellana S. , Hanene B. , Carluccio M.A. , Darra F. , Mate A. , Zimmermann A. , Gouider-Khouja N. and Valente E.M. , Mutations in B9D1 and MKS1 cause mild Joubert syndrome: Expanding the genetic overlap with the lethal ciliopathy Meckel syndrome, Orphanet J Rare Dis 9: ((2014) ), 72. |
[84] | Roosing S. , Romani M. , Isrie M. , Rosti R.O. , Micalizzi A. , Musaev D. , Mazza T. , Al-Gazali L. , Altunoglu U. , Boltshauser E. , D’Arrigo S. , De Keersmaecker B. , Kayserili H. , Brandenberger S. , Kraoua I. , Mark P.R. , McKanna T. , Van Keirsbilck J. , Moerman P. , Poretti A. , Puri R. , Van Esch H. , Gleeson J.G. and Valente E.M. , Mutations in CEP120 cause Joubert syndrome as well as complex ciliopathy phenotypes, J Med Genet 53: ((2016) ), 608–615. |
[85] | Roosing S. , Rosti R.O. , Rosti B. , de Vrieze E. , Silhavy J.L. , van Wijk E. , Wakeling E. and Gleeson J.G. , Identification of a homozygous nonsense mutation in KIAAin a consanguineous family displaying Joubert syndrome, Hum Genet 135: ((2016) ), 919–921. |
[86] | Sanders A.A. , de Vrieze E. , Alazami A.M. , Alzahrani F. , Alzahrani F. , Malarkey E.B. , Sorusch N. , Tebbe L. , Kuhns S. , van Dam T.J. , Alhashem A. , Tabarki B. , Lu Q. , Lambacher N.J. , Kennedy J.E. , Bowie R.V. , Hetterschijt L. , van Beersum S. , van Reeuwijk J. , Boldt K. , Kremer H. , Kesterson R.A. , Monies D. , Abouelhoda M. , Roepman R. , Huynen M.H. , Ueffing M. , Russell R.B. , Wolfrum U. , Yoder B.K. , van Wijk E. , Alkuraya F.S. and Blacque O.E. , KIAAis a novel ciliary basal body component mutated in Joubert syndrome, Genome Biol 16: ((2015) ), 293. |
[87] | Sang L. , Miller J.J. , Corbit K.C. , Giles R.H. , Brauer M.J. , Otto E.A. , Baye L.M. , Wen X. , Scales S.J. , Kwong M. , Huntzicker E.G. , Sfakianos M.K. , Sandoval W. , Bazan J.F. , Kulkarni P. , Garcia-Gonzalo F.R. , Seol A.D. , O’Toole J.F. , Held S. , Reutter H.M. , Lane W.S. , Rafiq M.A. , Noor A. , Ansar M. , Devi A.R. , Sheffield V.C. , Slusarski D.C. , Vincent J.B. , Doherty D.A. , Hildebrandt F. , Reiter J.F. and Jackson P.K. , Mapping the NPHP-JBTS-MKS protein network reveals ciliopathy disease genes and pathways, Cell 145: ((2011) ), 513–528. |
[88] | Satran D. , Pierpont M.E. and Dobyns W.B. , Cerebello-oculo-renal syndromes including Arima, Senior-Loken and COACH syndromes: More than just variants of Joubert syndrome, Am J Med Genet 86: ((1999) ), 459–469. |
[89] | Saunier S. , Calado J. , Benessy F. , Silbermann F. , Heilig R. , Weissenbach J. and Antignac C. , Characterization of the NPHP1 locus: Mutational mechanism involved in deletions in familial juvenile nephronophthisis, Am J Hum Genet 66: ((2000) ), 778–789. |
[90] | Sayer J.A. , Otto E.A. , O’Toole J.F. , Nurnberg G. , Kennedy M.A. , Becker C. , Hennies H.C. , Helou J. , Attanasio M. , Fausett B.V. , Utsch B. , Khanna H. , Liu Y. , Drummond I. , Kawakami I. , Kusakabe T. , Tsuda M. , Ma L. , Lee H. , Larson R.G. , Allen S.J. , Wilkinson C.J. , Nigg E.A. , Shou C. , Lillo C. , Williams D.S. , Hoppe B. , Kemper M.J. , Neuhaus T. , Parisi M.A. , Glass I.A. , Petry M. , Kispert A. , Gloy J. , Ganner A. , Walz G. , Zhu X. , Goldman D. , Nurnberg P. , Swaroop A. , Leroux M.R. and Hildebrandt F. , The centrosomal protein nephrocystin-6 is mutated in Joubert syndrome and activates transcription factor ATF4, Nat Genet 38: ((2006) ), 674–681. |
[91] | Schmidts M. , Arts H.H. , Bongers E.M. , Yap Z. , Oud M.M. , Antony D. , Duijkers L. , Emes R.D. , Stalker J. , Yntema J.B. , Plagnol V. , Hoischen A. , Gilissen C. , Forsythe E. , Lausch E. , Veltman J.A. , Roeleveld N. , Superti-Furga A. , Kutkowska-Kazmierczak A. , Kamsteeg E.J. , Elcioglu N. , van Maarle M.C. , Graul-Neumann L.M. , Devriendt K. , Smithson S.F. , Wellesley D. , Verbeek N.E. , Hennekam R.C. , Kayserili H. , Scambler P.J. , Beales P.L. , Uk10K, N.V. Knoers, R. Roepman and H.M. Mitchison, Exome sequencing identifies DYNC2H1 mutations as a common cause of asphyxiating thoracic dystrophy (Jeune syndrome) without major polydactyly, renal or retinal involvement, J Med Genet 50: ((2013) ), 309–323. |
[92] | Senocak E.U. , Oguz K.K. , Haliloglu G. , Topcu M. and Cila A. , Structural abnormalities of the brain other than molar tooth sign in Joubert syndrome-related disorders, Diagn Interv Radiol 16: ((2010) ), 3–6. |
[93] | Shaheen R. , Almoisheer A. , Faqeih E. , Babay Z. , Monies D. , Tassan N. , Abouelhoda M. , Kurdi W. , Al Mardawi E. , Khalil M.M. , Seidahmed M.Z. , Alnemer M. , Alsahan N. , Sogaty S. , Alhashem A. , Singh A. , Goyal M. , Kapoor S. , Alomar R. , Ibrahim N. , Alkuraya F.S. , Identification of a novel MKS locus defined by TMEM107 mutation, Hum Mol Genet 24: ((2015) ), 5211–5218. |
[94] | Shaheen R. , Faqeih E. , Seidahmed M.Z. , Sunker A. , Alali F.E. , AlQahtani K. and Alkuraya F.S. , A TCTN2 mutation defines a novel Meckel Gruber syndrome locus, Hum Mutat 32: ((2011) ), 573–578. |
[95] | Shaheen R. , Schmidts M. , Faqeih E. , Hashem A. , Lausch E. , Holder I. , Superti-Furga A. , Consortium U.K. , Mitchison H.M. , Almoisheer A. , Alamro R. , Alshiddi T. , Alzahrani F. , Beales P.L. and Alkuraya F.S. , A founder CEP120 mutation in Jeune asphyxiating thoracic dystrophy expands the role of centriolar proteins in skeletal ciliopathies, Hum Mol Genet 24: ((2015) ), 1410–1419. |
[96] | Shaheen R. , Shamseldin H.E. , Loucks C.M. , Seidahmed M.Z. , Ansari S. , Ibrahim Khalil M. , Al-Yacoub N. , Davis E.E. , Mola N.A. , Szymanska K. , Herridge W. , Chudley A.E. , Chodirker B.N. , Schwartzentruber J. , Majewski J. , Katsanis N. , Poizat C. , Johnson C.A. , Parboosingh J. , Boycott K.M. , Innes A.M. , Alkuraya F.S. , Mutations in CSPP1, encoding a core centrosomal protein, cause a range of ciliopathy phenotypes in humans, Am J Hum Genet 94: ((2014) ), 73–79. |
[97] | Shaheen R. , Szymanska K. , Basu B. , Patel N. , Ewida N. , Faqeih E. , Al Hashem A. , Derar N. , Alsharif H. , Aldahmesh M.A. , Alazami A.M. , Hashem M. , Ibrahim N. , Abdulwahab F.M. , Sonbul R. , Alkuraya H. , Alnemer M. , Al Tala S. , Al-Husain M. , Morsy H. , Seidahmed M.Z. , Meriki N. , Al-Owain M. , AlShahwan S. , Tabarki B. , Salih M.A. , Ciliopathy W. , Faquih T. , El-Kalioby M. , Ueffing M. , Boldt K. , Logan C.V. , Parry D.A. , Al Tassan N. , Monies D. , Megarbane A. , Abouelhoda M. , Halees A. , Johnson C.A. , Alkuraya F.S. , Characterizing the morbid genome of ciliopathies, Genome Biol 17: ((2016) ), 242. |
[98] | Slaats G.G. , Isabella C.R. , Kroes H.Y. , Dempsey J.C. , Gremmels H. , Monroe G.R. , Phelps I.G. , Duran K.J. , Adkins J. , Kumar S.A. , Knutzen D.M. , Knoers N.V. , Mendelsohn N.J. , Neubauer D. , Mastroyianni S.D. , Vogt J. , Worgan L. , Karp N. , Bowdin S. , Glass I.A. , Parisi M.A. , Otto E.A. , Johnson C.A. , Hildebrandt F. , van Haaften G. , Giles R.H. and Doherty D. , MKS1 regulates ciliary INPP5E levels in Joubert syndrome, J Med Genet 53: ((2016) ), 62–72. |
[99] | Srivastava S. , Molinari E. , Raman S. and Sayer J.A. , Many Genes-One Disease? Genetics of Nephronophthisis (NPHP) and NPHP-Associated Disorders, Front Pediatr 5: ((2017) ), 287. |
[100] | Srour M. , Hamdan F.F. , McKnight D. , Davis E. , Mandel H. , Schwartzentruber J. , Martin B. , Patry L. , Nassif C. , Dionne-Laporte A. , Ospina L.H. , Lemyre E. , Massicotte C. , Laframboise R. , Maranda B. , Labuda D. , Decarie J.C. , Rypens F. , Goldsher D. , Fallet-Bianco C. , Soucy J.F. , Laberge A.M. , Maftei C. , Care4Rare Canada C. , Boycott K. , Brais B. , Boucher R.M. , Rouleau G.A. , Katsanis N. , Majewski J. , Elpeleg O. , Kukolich M.K. , Shalev S. and Michaud J.L. Joubert syndrome in french canadians and identification of mutations in CEP104, Am J Hum Genet 97: ((2015) ), 744–753. |
[101] | Srour M. , Hamdan F.F. , Schwartzentruber J.A. , Patry L. , Ospina L.H. , Shevell M.I. , Desilets V. , Dobrzeniecka S. , Mathonnet G. , Lemyre E. , Massicotte C. , Labuda D. , Amrom D. , Andermann E. , Sebire G. , Maranda B. , Consortium F.C. , Rouleau G.A. , Majewski J. and Michaud J.L. , Mutations in TMEM231 cause Joubert syndrome in French Canadians, J Med Genet 49: ((2012) ), 636–641. |
[102] | Srour M. , Schwartzentruber J. , Hamdan F.F. , Ospina L.H. , Patry L. , Labuda D. , Massicotte C. , Dobrzeniecka S. , Capo-Chichi J.M. , Papillon-Cavanagh S. , Samuels M.E. , Boycott K.M. , Shevell M.I. , Laframboise R. , Desilets V. , Consortium F.C. , Maranda B. , Rouleau G.A. , Majewski J. and Michaud J.L. , Mutations in C5ORF42 cause Joubert syndrome in the French Canadian population, Am J Hum Genet 90: ((2012) ), 693–700. |
[103] | Steinlin M. , Schmid M. , Landau K. and Boltshauser E. , Follow-up in children with Joubert syndrome, Neuropediatrics 28: ((1997) ), 204–211. |
[104] | Strongin A. , Heller T. , Doherty D. , Glass I.A. , Parisi M.A. , Bryant J. , Choyke P. , Turkbey B. , Daryanani K. , Yildirimli D. , Vemulapalli M. , Mullikin J.C. , Malicdan M.C. , Vilboux T. , Gahl W.A. , Gunay-Aygun M. and Program N.C.S. , Characteristics of liver disease in 100 individuals with joubert syndrome prospectively evaluated at a single center, J Pediatr Gastroenterol Nutr 66: ((2018) ), 428–435. |
[105] | Summers A.C. , Snow J. , Wiggs E. , Liu A.G. , Toro C. , Poretti A. , Zein W.M. , Brooks B.P. , Parisi M.A. , Inati S. , Doherty D. , Vemulapalli M. , Mullikin J.C. , Program N.C.S. , Vilboux T. , Gahl W.A. and Gunay-Aygun M. , Neuropsychological phenotypes of 76 individuals with Joubert syndrome evaluated at a single center, Am J Med Genet A ((2017) ). |
[106] | Suzuki T. , Miyake N. , Tsurusaki Y. , Okamoto N. , Alkindy A. , Inaba A. , Sato M. , Ito S. , Muramatsu K. , Kimura S. , Ieda D. , Saitoh S. , Hiyane M. , Suzumura H. , Yagyu K. , Shiraishi H. , Nakajima M. , Fueki N. , Habata Y. , Ueda Y. , Komatsu Y. , Yan K. , Shimoda K. , Shitara Y. , Mizuno S. , Ichinomiya K. , Sameshima K. , Tsuyusaki Y. , Kurosawa K. , Sakai Y. , Haginoya K. , Kobayashi Y. , Yoshizawa C. , Hisano M. , Nakashima M. , Saitsu H. , Takeda S. and Matsumoto N. , Molecular genetic analysis of 30 families with Joubert syndrome, Clin Genet 90: ((2016) ), 526–535. |
[107] | Thomas S. , Cantagrel V. , Mariani L. , Serre V. , Lee J.E. , Elkhartoufi N. , de Lonlay P. , Desguerre I. , Munnich A. , Boddaert N. , Lyonnet S. , Vekemans M. , Lisgo S.N. , Caspary T. , Gleeson J. and Attie-Bitach T. , Identification of a novel ARL13B variant in a Joubert syndrome-affected patient with retinal impairment and obesity, Eur J Hum Genet ((2014) ). |
[108] | Thomas S. , Legendre M. , Saunier S. , Bessieres B. , Alby C. , Bonniere M. , Toutain A. , Loeuillet L. , Szymanska K. , Jossic F. , Gaillard D. , Yacoubi M.T. , Mougou-Zerelli S. , David A. , Barthez M.A. , Ville Y. , Bole-Feysot C. , Nitschke P. , Lyonnet S. , Munnich A. , Johnson C.A. , Encha-Razavi F. , Cormier-Daire V. , Thauvin-Robinet C. , Vekemans M. and Attie-Bitach T. , TCTN3 mutations cause Mohr-Majewski syndrome, Am J Hum Genet 91: ((2012) ), 372–378. |
[109] | Thomas S. , Wright K.J. , Le Corre S. , Micalizzi A. , Romani M. , Abhyankar A. , Saada J. , Perrault I. , Amiel J. , Litzler J. , Filhol E. , Elkhartoufi N. , Kwong M. , Casanova J.L. , Boddaert N. , Baehr W. , Lyonnet S. , Munnich A. , Burglen L. , Chassaing N. , Encha-Ravazi F. , Vekemans M. , Gleeson J.G. , Valente E.M. , Jackson P.K. , Drummond I.A. , Saunier S. and Attie-Bitach T. , A homozygous PDE6D mutation in Joubert syndrome impairs targeting of farnesylated INPP5E protein to the primary cilium, Hum Mutat 35: ((2014) ), 137–146. |
[110] | Toriello H.V. , Franco B. , Bruel A.-L. and Thauvin-Robinet C. , Oral-Facial-Digital Syndrome Type I, 2002 Jul 24 [Updated 2016 Aug 4]. In: M.P. Adam, H.H. Ardinger, R.A. Pagon, et al., editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993–2019. |
[111] | Tory K. , Lacoste T. , Burglen L. , Moriniere V. , Boddaert N. , Macher M.A. , Llanas B. , Nivet H. , Bensman A. , Niaudet P. , Antignac C. , Salomon R. and Saunier S. , High NPHP1 and NPHP6 mutation rate in patients with Joubert syndrome and nephronophthisis: Potential epistatic effect of NPHP6 and AHI1 mutations in patients with NPHP1 mutations, J Am Soc Nephrol 18: ((2007) ), 1566–1575. |
[112] | Tuz K. , Bachmann-Gagescu R. , O’Day D.R. , Hua K. , Isabella C.R. , Phelps I.G. , Stolarski A.E. , O’Roak B.J. , Dempsey J.C. , Lourenco C. , Alswaid A. , Bonnemann C.G. , Medne L. , Nampoothiri S. , Stark Z. , Leventer R.J. , Topcu M. , Cansu A. , Jagadeesh S. , Done S. , Ishak G.E. , Glass I.A. , Shendure J. , Neuhauss S.C. , Haldeman-Englert C.R. , Doherty D. and Ferland R.J. , Mutations in CSPP1 cause primary cilia abnormalities and Joubert syndrome with or without Jeune asphyxiating thoracic dystrophy, Am J Hum Genet 94: ((2014) ), 62–72. |
[113] | Utsch B. , Sayer J.A. , Attanasio M. , Pereira R.R. , Eccles M. , Hennies H.C. , Otto E.A. and Hildebrandt F. , Identification of the first AHI1 gene mutations in nephronophthisis-associated Joubert syndrome, Pediatr Nephrol 21: ((2006) ), 32–35. |
[114] | Valente E.M. , Logan C.V. , Mougou-Zerelli S. , Lee J.H. , Silhavy J.L. , Brancati F. , Iannicelli M. , Travaglini L. , Romani S. , Illi B. , Adams M. , Szymanska K. , Mazzotta A. , Lee J.E. , Tolentino J.C. , Swistun D. , Salpietro C.D. , Fede C. , Gabriel S. , Russ C. , Cibulskis K. , Sougnez C. , Hildebrandt F. , Otto E.A. , Held S. , Diplas B.H. , Davis E.E. , Mikula M. , Strom C.M. , Ben-Zeev B. , Lev D. , Sagie T.L. , Michelson M. , Yaron Y. , Krause A. , Boltshauser E. , Elkhartoufi N. , Roume J. , Shalev S. , Munnich A. , Saunier S. , Inglehearn C. , Saad A. , Alkindy A. , Thomas S. , Vekemans M. , Dallapiccola B. , Katsanis N. , Johnson C.A. , Attie-Bitach T. and Gleeson J.G. , Mutations in TMEM216 perturb ciliogenesis and cause Joubert, Meckel and related syndromes, Nat Genet 42: ((2010) ), 619–625. |
[115] | Valente E.M. , Silhavy J.L. , Brancati F. , Barrano G. , Krishnaswami S.R. , Castori M. , Lancaster M.A. , Boltshauser E. , Boccone L. , Al-Gazali L. , Fazzi E. , Signorini S. , Louie C.M. , Bellacchio E. , International Joubert Syndrome Related Disorders Study G. , Bertini E. , Dallapiccola B. and Gleeson J.G. , Mutations in CEP290, which encodes a centrosomal protein, cause pleiotropic forms of Joubert syndrome, Nat Genet 38: ((2006) ), 623–625. |
[116] | Van De Weghe J.C. , Rusterholz T.D.S. , Latour B. , Grout M.E. , Aldinger K.A. , Shaheen R. , Dempsey J.C. , Maddirevula S. , Cheng Y.H. , Phelps I.G. , Gesemann M. , Goel H. , Birk O.S. , Alanzi T. , Rawashdeh R. , Khan A.O. , University of Washington Center for Mendelian G. , Bamshad M.J. , Nickerson D.A. , Neuhauss S.C.F. , Dobyns W.B. , Alkuraya F.S. , Roepman R. , Bachmann-Gagescu R. and Doherty D. , Mutations in ARMC9, which encodes a basal body protein, cause joubert syndrome in humans and ciliopathy phenotypes in zebrafish, Am J Hum Genet 101: ((2017) ), 23–36. |
[117] | Verloes A. and Lambotte C. , Further delineation of a syndrome of cerebellar vermis hypo/aplasia, oligophrenia, congenital ataxia, coloboma, and hepatic fibrosis, Am J Med Genet 32: ((1989) ), 227–232. |
[118] | Vilboux T. , Doherty D.A. , Glass I.A. , Parisi M.A. , Phelps I.G. , Cullinane A.R. , Zein W. , Brooks B.P. , Heller T. , Soldatos A. , Oden N.L. , Yildirimli D. , Vemulapalli M. , Mullikin J.C. and Nisc Comparative Sequencing P. , Malicdan M.C. , Gahl W.A. and Gunay-Aygun M. , Molecular genetic findings and clinical correlations in 100 patients with Joubert syndrome and related disorders prospectively evaluated at a single center, Genet Med ((2017) ). |
[119] | Wang S.F. , Kowal T.J. , Ning K. , Koo E.B. , Wu A.Y. , Mahajan V.B. and Sun Y. , Review of Ocular Manifestations of Joubert Syndrome, Genes (Basel) 9: ((2018) ). |
[120] | Wheway G. , Schmidts M. , Mans D.A. , Szymanska K. , Nguyen T.T. , Racher H. , Phelps I.G. , Toedt G. , Kennedy J. , Wunderlich K.A. , Sorusch N. , Abdelhamed Z.A. , Natarajan S. , Herridge W. , van Reeuwijk J. , Horn N. , Boldt K. , Parry D.A. , Letteboer S.J.F. , Roosing S. , Adams M. , Bell S.M. , Bond J. , Higgins J. , Morrison E.E. , Tomlinson D.C. , Slaats G.G. , van Dam T.J.P. , Huang L. , Kessler K. , Giessl A. , Logan C.V. , Boyle E.A. , Shendure J. , Anazi S. , Aldahmesh M. , Al Hazzaa S. , Hegele R.A. , Ober C. , Frosk P. , Mhanni A.A. , Chodirker B.N. , Chudley A.E. , Lamont R. , Bernier F.P. , Beaulieu C.L. , Gordon P. , Pon R.T. , Donahue C. , Barkovich A.J. , Wolf L. , Toomes C. , Thiel C.T. , Boycott K.M. , McKibbin M. , Inglehearn C.F. , Consortium U.K. , University of Washington Center for Mendelian G. , Stewart F. , Omran H. , Huynen M.A. , Sergouniotis P.I. , Alkuraya F.S. , Parboosingh J.S. , Innes A.M. , Willoughby C.E. , Giles R.H. , Webster A.R. , Ueffing M. , Blacque O. , Gleeson J.G. , Wolfrum U. , Beales P.L. , Gibson T. , Doherty D. , Mitchison H.M. , Roepman R. and Johnson C.A. , An siRNA-based functional genomics screen for the identification of regulators of ciliogenesis and ciliopathy genes, Nat Cell Biol 17: ((2015) ), 1074–1087. |
[121] | Wolf M.T. , Saunier S. , O’Toole J.F. , Wanner N. , Groshong T. , Attanasio M. , Salomon R. , Stallmach T. , Sayer J.A. , Waldherr R. , Griebel M. , Oh J. , Neuhaus T.J. , Josefiak U. , Antignac C. , Otto E.A. and Hildebrandt F. , Mutational analysis of the RPGRIP1L gene in patients with Joubert syndrome and nephronophthisis, Kidney Int 72: ((2007) ), 1520–1526. |


