
Blueberries and insulin protect microglial cells against high glucose-induced inflammation and restore GLUT-1
Abstract
BACKGROUND:
Growing evidence suggests that hyperglycemia could be harmful for cognitive function. That insulin (INS) has a neuro-modulatory role is supported by various findings, but its effect on microglia, the innate immune cells in the brain, is largely unknown. Blueberries have been shown to reduce neuro-inflammation.
OBJECTIVE:
We hypothesized that high glucose stimulated an inflammation in microglia and that BB and INS were able to reduce it and both might act through GLUT-1 transporter.
METHODS:
We examined the effects of low (5 mM), medium (25 mM), or high (50 mM) glucose, stimulated or not with lipopolysaccharide (LPS; 100 nM) with either BB extract (2 mg/ml) and/or INS, on inflammatory responses in a microglia cell line. Nitric oxide (NO) production and the expression levels of iNOS, TNF-α, NOX4 and glucose transporter protein-1 (GLUT1) were assessed.
RESULTS:
We observed that treatment with BB, similarly to INS treatments, reduced the high glucose concentration-induced response on oxidative stress and inflammation, and that this protective effect is more important with LPS added to glucose media. Interestingly, both BB and INS attenuated the LPS-induced inflammatory response on GLUT1.
CONCLUSION:
Increasing glucose concentration triggers inflammation by microglia. BB as well as INS protected microglia from high glucose levels, by reducing inflammation and altering glucose transport in microglia. These preliminary data compared for the first time BB to Insulin on microglia. Blueberries are promising dietary intervention to prevent diabetic neuropathy. Our preliminary results suggest a possible new mechanism involving GLUT-1 by which BB has insulin-like effects.
Abbreviations
BB | Blueberries |
CNS | Central nervous system |
GLUT | Glucose transporter |
HAPI | Highly Aggressively Proliferating Immortalized |
INS | Insulin |
NFκB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
NOX | NADPH oxidase |
NOS | Nitric oxide synthases |
Nrf2 | NFE-E2-related factor 2 |
LPS | Lipopolysaccharide |
TNF-α | Tumor necrosis factor-alpha |
ROS | Reactive oxygen species |
1Introduction
Growing evidence suggests that hyperglycemia could be damaging for cognitive function [1]. Therefore, with the rising prevalence of the aging population, as well as those with type 2 diabetes which steadily increases with age, neurodegenerative diseases are projected to rise dramatically worldwide [1]. Clinical and animal studies have linked high blood glucose and chronic uncontrolled diabetes mellitus to memory impairment and cognitive dysfunction, thus suggesting that hyperglycemia may contribute to learning and memory decline [2]. Uncontrolled prolonged hyperglycemia can lead to excessive amounts of reactive oxygen species (ROS), which trigger inflammation, leading to impairment in brain functions [3]. So far, activation of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) and inducible nitric oxide synthases (iNOS), leading to an overproduction of ROS, is the main robust hypothesis for linking hyperglycemia to oxidative stress [4]. New information from in vivo and in vitro studies support the evidence that insulin plays a key physiological role in the brain. Besides regulating peripheral glucose homeostasis, insulin is an important neuromodulator that contributes to neurobiological processes, while insulin resistance may contribute to cognitive deficits in aging and Alzheimer disease (AD) [5–7]. However, the relevance of its molecular effects remains to be elucidated.
Microglia are the resident immune cells playing major roles in the central nervous system (CNS) in both health and disease [8]. Although microglial activation is a necessary and beneficial response to CNS insults, uncontrolled microglial activation contributes critically to aging and to the pathogenesis of various neurodegenerative diseases [9] such as diabetic neuropathy [10] and AD [11]. Pro-inflammatory microglia release excessive amounts of nitric oxide (NO) and ROS such as superoxide [9]. NOX has been identified as a major contributor of superoxide production in microglia [12, 13] and has been involved in cerebral ischemia and neurodegenerative disease [9–11]. Therefore, targeting NOX in microglia might lead to therapeutic opportunities.
Even though chronic uncontrolled hyperglycemia is associated with inflammation at the systemic level, there are few studies examining the effect of high glucose and INS on microglia. It has been demonstrated that microglia express several glucose transporters, in particularly GLUT1, in rodents to guarantee sufficient glucose influx for their energy requirement [14]. Interestingly, in a mouse model of AD, significant reduction in microglia activation was observed following intranasal insulin (INS) treatment [15] suggesting anti-inflammatory effect of Insulin.
Human and experimental studies assessing the effects of foods high in polyphenols, such as anthocyanin-rich blueberries (BBs), have revealed promising findings indicating that they prevent cognitive decline [16, 17]. Previous studies from our laboratory have shown that dietary intake of BB improves memory, attenuates oxidative stress, and reduces inflammatory signals [18], as well as improves task-related activation in healthy older adults [19]. BBs can modulate a variety of biochemical/signaling pathways that promote cognitive enhancement, which has created interest in their mechanisms of action [20]. It is noteworthy that in population studies in individuals with prediabetes and type 2 diabetes, of all the polyphenols analyzed, only BB and anthocyanin intake was associated with improvements in insulin resistance and a decrease in inflammation [21]. This benefit has been recently confirmed in a double-blind, randomized study [22]. Indeed, classical antioxidant actions of polyphenols undoubtedly contribute to their anti-inflammatory roles by interrupting reactive oxygen species (ROS) as an inducer of inflammation through the stimulation of NF-kB. NF-kB is a key regulator in innate immune/inflammatory stress responses, triggering the expression of a large numbers of genes including various inflammatory cytokines such as COX-2, NO synthase, and TNF-α [23]. The role of polyphenols as Nrf2 inducers is a unifying mechanism by which polyphenols exert both antioxidant and anti-inflammatory properties, because Nrf2 triggers an antioxidant response leading to an inhibitory ROS decrease to trigger NfκB [24], taking a part in their beneficial role in the prevention and the progress of chronic diseases related to inflammation [25]. Berries are unique because they are rich sources of anthocyanin polyphenols and data are consistent to support that anthocyanins are an important part of the biological activity delivered by berries to modulate inflammation [26, 27]. Emerging interest is also focused on the potential of berries such as strawberry, not only to activate AMPK, but to possibly alter mitochondrial function, leading to amelioration of insulin sensitivity [28, 29]. One other possible way might be via upregulation of GLUT gene expression [30]. However, little is known about the neurobiological effect of action of BBs on hyperglycemia in the brain, and the key regulating proteins involved in neuroinflammation under varying glycemic conditions.
In an effort to better understand the role of BBs on the CNS, we hypothesized that hyperglycemia would up-regulate inflammatory mediators, which would be dampened by the phytochemicals found in BBs, which might act by an INS-like effect. Therefore, we investigated whether microglia exposed to a high glucose media would show increased expression of neuroinflammatory mediators, with and without lipopolysaccharide (LPS) stimulation. LPS-activated microglia provide an in vitro model for the study of mechanisms underlying inflammation-mediated neuronal damage [31]. We then investigated whether insulin and blueberries were able to counteract the induced high-glucose effects in both environments and whether they modulated GLUT-1 expression, since glucose uptake in rat microglia is dominated by GLUT-1 [14].
2Methods
2.1Cell culture
HAPI rat microglia cells (generously provided by Dr. Grace Sun, University of Missouri, Columbia MS) were plated on 12-well plates for immunocytochemical analysis at a density of 50,000 cells per well or on 96-well plates for the enzyme-linked immunosorbent assay (ELISA) at a density of 10,000 cells per well. The cells were maintained in low (5 mM), normal (25 mM), or high (50 mM) glucose in Dulbecco’s modified Eagle’s medium (DMEM, Invitrogen, Carlsbad, CA) supplemented with 10% fetal calf serum, 100 U/mL penicillin, and 100 mg/mL streptomycin (Invitrogen, Carlsbad, CA) at 37°C in a humidified incubator under 5% CO2. The glucose levels were determined based on Routh et al. [32], who reported that physiological hypothalamic glucose level varies from ∼0.7 to 2.5 mM between the fasted and fed states. However, numerous in vitro studies investigating glucose-sensing neurons still use standard extracellular solutions containing 10–20 mM glucose, which is similar to blood glucose levels occurring during diabetes. When the cells were nearly 70% confluent, they were pre-treated with either insulin (50 mM, Sigma-Aldrich, MO) or BB (2 mg/ml) or with a combination of INS and BB for 24 hours. BB was prepared and administered as described elsewhere [33]. Then, cells were stimulated with LPS (100 ng/mL, Sigma-Aldrich, MO) overnight, to evaluate the impact of glucose level on the LPS-induced inflammation and the protective effects of BB and INS to prevent LPS inflammation.
2.2Nitrite quantification
The extracellular release of nitrite (NO2-) by HAPI cells under various treatment conditions was measured by Griess reagent (Promega, Madison, WI) according to the manufacturer’s instructions. 100 ml of cell-conditioned media supernatant was mixed with 25 ml of Griess reagent (0.1% N-1-naphthylethylenediamine dihydrochloride and 1% sulphanilamide in 5% phosphoric acid) in a 96-well plate and incubated at room temperature for 20 minutes. Absorbance was read at 548 nm and the concentration of nitrite was calculated with the linear equation derived from the standard curve generated by known concentrations of sodium nitrite.
2.3TNF-α ELISA
Quantification of tumor necrosis factor-alpha (TNF-α) in cell-conditioned media was performed with an enzyme-linked immunosorbent assay (ELISA, eBioscience, San Diego, CA) according to the manufacturer’s instructions. TNF-α concentration for each sample was calculated from the linear equation derived from the standard curve of known concentrations of the cytokine.
2.4Western blots
Western blots were performed as described in Poulose et al. [34]. In brief, after the treatments, cells were lysed at 4°C with CelLytic-M Cell Lysis Reagent (Sigma-Aldrich, St. Louis, MO), containing protease inhibitor cocktail (Roche, Basel, Switzerland), PMSF (10 μg/mL), and sodium fluoride (50 mM) (Sigma-Aldrich), centrifuged at 12,000 g for 15 min at 4°C, and protein concentration was quantified with the DC protein assay (Bio-Rad, Hercules, CA). Equal amounts of denatured protein samples were separated by 8% SDS-PAGE, electrophoretically transferred to nitrocellulose membrane, and probed with primary antibodies for iNOS (Upstate-Millipore, Billerica, MA). After overnight incubation, the blots were incubated with the appropriate HRP-conjugated secondary antibody (Upstate-Millipore). The primary antibodies were used at 1:1000 dilutions. Secondary antibodies were used at 1:4000 dilutions. The signal was detected using an Enhanced Chemiluminescence (ECL) detection kit (Amersham Corporation, Arlington Heights, IL). The blots were imaged (EC3 Darkroom, UVP, Upland CA) and band densities were analyzed using LabWorks Imaging Acquisition and Analysis software (version 4.5, UVP). β-actin was used as a loading control.
2.5Immunofluorescence staining
Fluorescent imaging for the accumulation of the expression of glucose transporter GLUT1 was done as described previously [35]. Briefly, cells at 30% confluence were grown in 8-well chamber slides (Menzel-Gläser) with different glucose conditions (Low, Medium, and High), and then were pretreated 24 hours with respective concentrations of BB extracts and/or INS. The media was removed after the pretreatment period, and cells were stimulated with LPS for 2 hours. After the treatment, cells were washed with PBS, fixed in 100% cold methanol for 10 min, and washed 3 times with PBS. The slides were blocked in 5% normal goat serum prepared in 0.1% PBS containing 0.1% Triton-100. The slides were incubated overnight at 4°C in NOX4 (NOX4/ab133303, Abcam, Cambridge, MA) or GLUT1 antibody (GLUT1/ab652, Abcam, Cambridge, MA) at a 1:500 dilution of primary antibody, and after removal of antibody, cells were washed with PBS and incubated in TRITC or FITC -conjugated secondary antibody (Cell Signaling, Danvers, MA). The secondary antibody was removed, and slides were washed thoroughly with PBS 3 times with minimal light exposure and mounted with glycerol-based Antifade (Sigma-Aldrich, St. Louis, MO) containing DAPI. The images were taken using a Nikon Eclipse TE200U inverted fluorescence microscope coupled to a digital CCD camera (Hamamatsu Photonics, Bridgewater, NJ) and illuminated with a fluorescent light source. The images were analyzed using NIS-Elements Software (Nikon Instruments, Melville, NY).
2.6Statistical method
All statistical analyses were performed using SYSTAT software (Systat Software, Inc., San Jose, CA). Each experiment was repeated three times. Data were presented as mean ± SEM and analyzed using a two-way ANOVA or a three-way ANOVA when appropriate. Post-hoc comparisons were performed using Ducan or Fisher’s least significant difference (LSD) test, when either the main effect or interaction was significant. Results were considered statistically significant if the observed significance level with treatment was p < 0.05.
3Results
3.1Extracellular release of nitrite
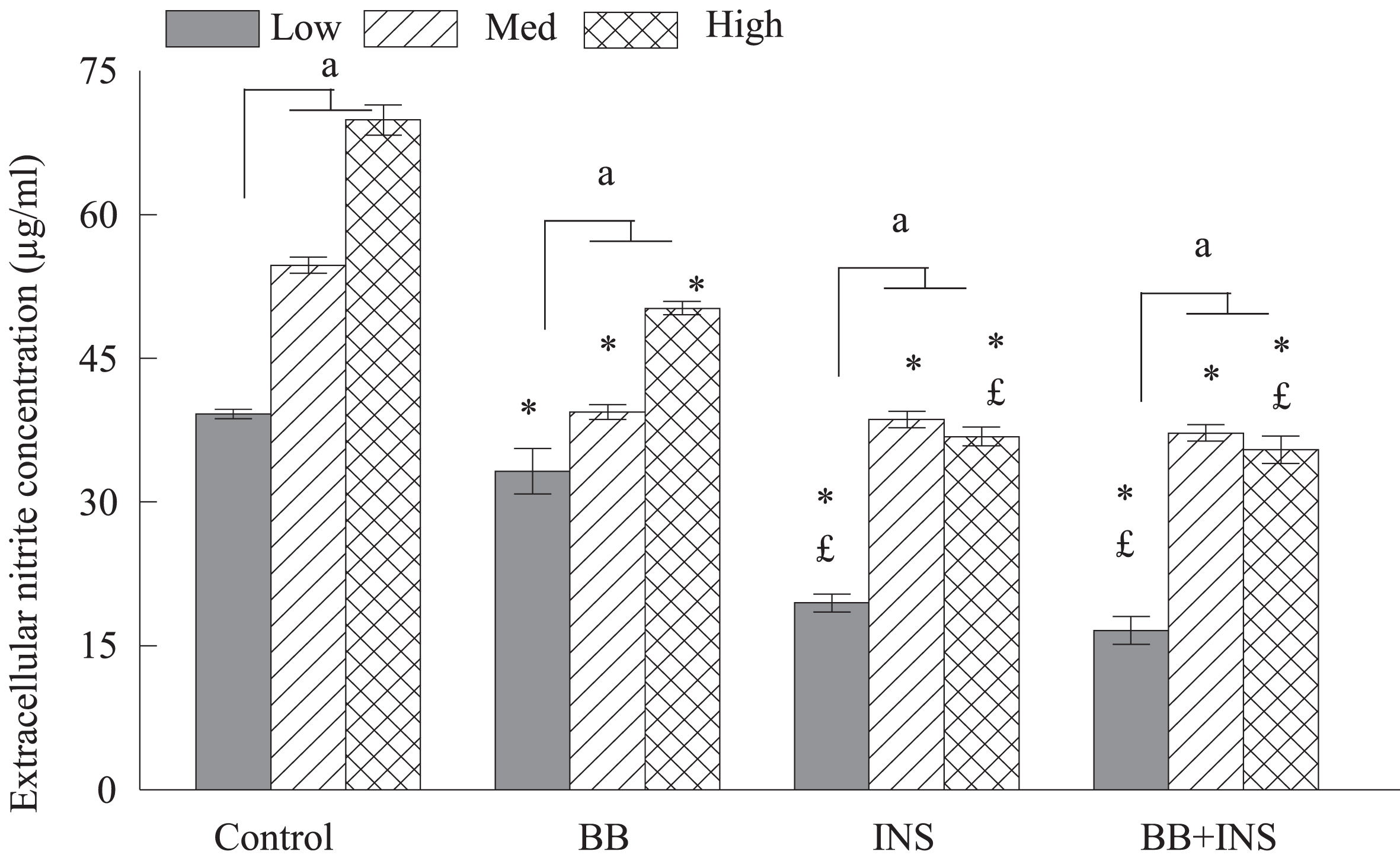
The release of nitrites by microglia under LPS stimulation is well-known but the effect of increasing glucose concentration on this production is not. Therefore, we assessed the nitrites production depending on three glucose concentrations (low, medium, high). The overall effect of glucose concentration following LPS treatment on extracellular nitrite production by HAPI cells was significant (F2,112 = 294.38, P < 0.0001) and Post-hoc comparisons indicated a significant proportional increase in nitrite levels with increasing glucose concentration (Fig. 1). The effects of each treatments BB and INS to counteract the LPS-induced nitrite production were also significant (BB: F1,112 = 114.67, P < 0.0001) and INS (INS: F1,112 = 553.20, P < 0.0001). Post-Hoc treatments confirmed that pre-treatments with BB (p < 0.05) and INS (P < 0.05) decrease significantly nitrite production compared to their respective glucose-matched LPS control (Fig. 1). Insulin was significantly more effective than BB at high glucose concentrations, but equivalent to BB at the medium glucose concentration. The combination of INS and BB had no additive effect when compared to INS alone (Fig. 1).
Fig. 1
Production of extracellular nitrite in HAPI microglial cells cultured in different concentrations of glucose (low, 5 mM; med, 25 mM; high, 50 mM) and pretreated with 2% blueberries (BB), 50 mM insulin (INS), or combination of blueberries and insulin for 4 hours. The cells were stimulated with LPS (100ng/mL) and extracellular levels of NO were determined using Greiss reagent. Data are expressed as mean±SEM and compared against control as LPS treatment alone in different glucose concentrations with levels of significance at p < 0.05. The letter and symbols mean statistical difference between treatments. “a” means statistical difference between medium or high glucose vs low glucose for each treatment; * means a statistical difference between treatments vs Control (LPS) matched for each glucose concentration; £ means a statistical difference between treatments BB vs INS and BB vs BB + INS. In all treatment condition (control; BB; Ins; Ins + BB) nitrite concentrations were significantly higher in medium and high glucose compared to low glucose (a < 0.01); medium and high glucose were not different for nitrite concentration (sharing the same letter a). Compared to control group, pretreatment with BB, Ins or Ins+BB decreased significantly (*< 0.05) nitrite concentration. The level of nitrite was significantly lower in Ins and Ins + BB compared to BB at low and high glucose concentration (£< 0.05).
3.2Expression of iNOS
Under basal condition (without LPS) the overall effect of glucose on the expression of iNOS was significant (F2,93 = 103.21, P < 0.0001), and with increasing concentration of glucose, the iNOS expression also substantially increased. The overall effect of blueberry on iNOS was not significant (P = 0.986) while INS had an overall attenuating effect on iNOS (F1,93 = 54.39, P < 0.0001). Post-hoc comparisons indicated that high glucose alone significantly increased the expression of iNOS as did BB treatment at medium and high glucose concentration, compared to their glucose-matched control (Fig. 2). However, both INS and BB plus INS combination significantly reduced the iNOS protein levels under high glucose conditions compared to their glucose-matched controls (Fig. 2).
Fig. 2
Expression levels of iNOS from microglial lysates cultured in various concentration of glucose and pretreated with 2% blueberries (BB), 50 mM insulin (INS), or combination of blueberries and insulin for 4 hours is expressed as the mean±SEM of three independent experiments with levels of significance at p < 0.05. a) Immunoblot probed with anti-iNOS (131kD) and actin (45kD) for low, medium and high glucose for control (glucose only), positive control (LPS), pretreated with BB or Ins b) Expression levels of iNOS as determined by the western blot immunoreactive band density, grown under the various concentrations of glucose in response to LPS and treatments with BB and INS. The letter or symbols mean a statistical difference between treatments. “a” means statistical difference between medium or high glucose vs low glucose for each treatments; *means a statistical difference between treatments vs Control respectively for each glucose concentration; # means a statistical difference between treatments vs LPS as a positive control respectively for each glucose concentration; £ means a statistical difference between BB vs others treatments (INS and INS+BB) respectively for each glucose concentration. Compared to low glucose levels of iNOS are significantly increased with high glucose in all conditions (control, BB Ins, Ins + BB without or with LPS) (a < 0.05). Insulin pretreatment significantly lowered iNOS expression compared to control (*< 0.05) at medium and high glucose. Under LPS stimulation, BB decreased significantly (# < 0.05) iNOS level. At high glucose + LPS condition, INS and INS + BB decrease significantly iNOS level compared to BB (£< 0.05).
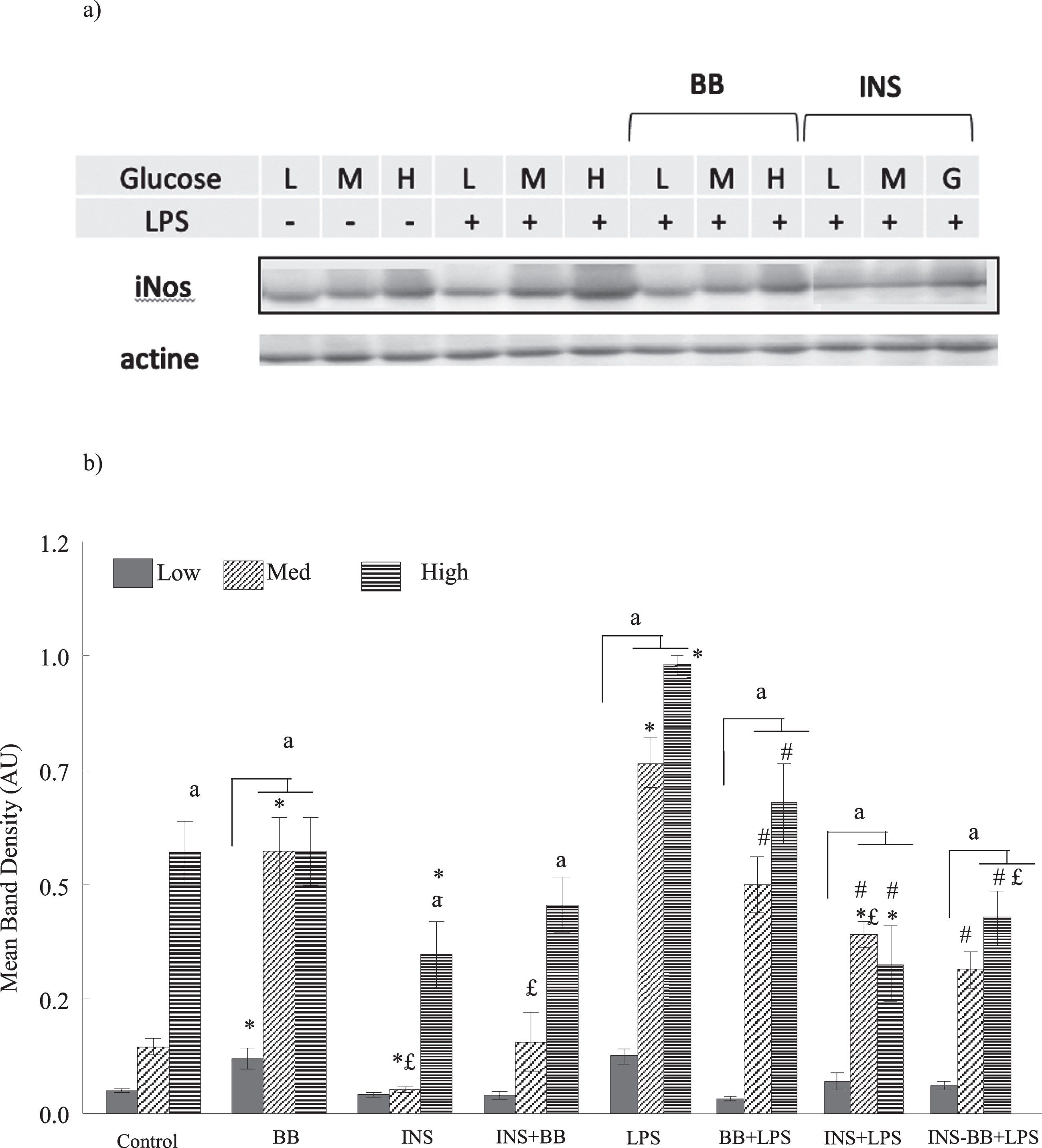
The main effect is LPS induced an overall significant increase in iNOS (F1,93 = 22.42, P < 0.0001). All interactions between LPS and treatments were significant and Post-hoc comparisons indicated a significant elevation in the levels of iNOS, under medium and high glucose conditions (p < 0.005) with LPS (Fig. 2). The pre-treatments with BB and INS alone or in combination (BB plus INS) significantly attenuated the iNOS expression compared to their glucose-matched LPS control (Fig. 2: *p < 0.005), indicating BB was more effective under LPS-induced stress conditions.
3.3Expression of TNF- α
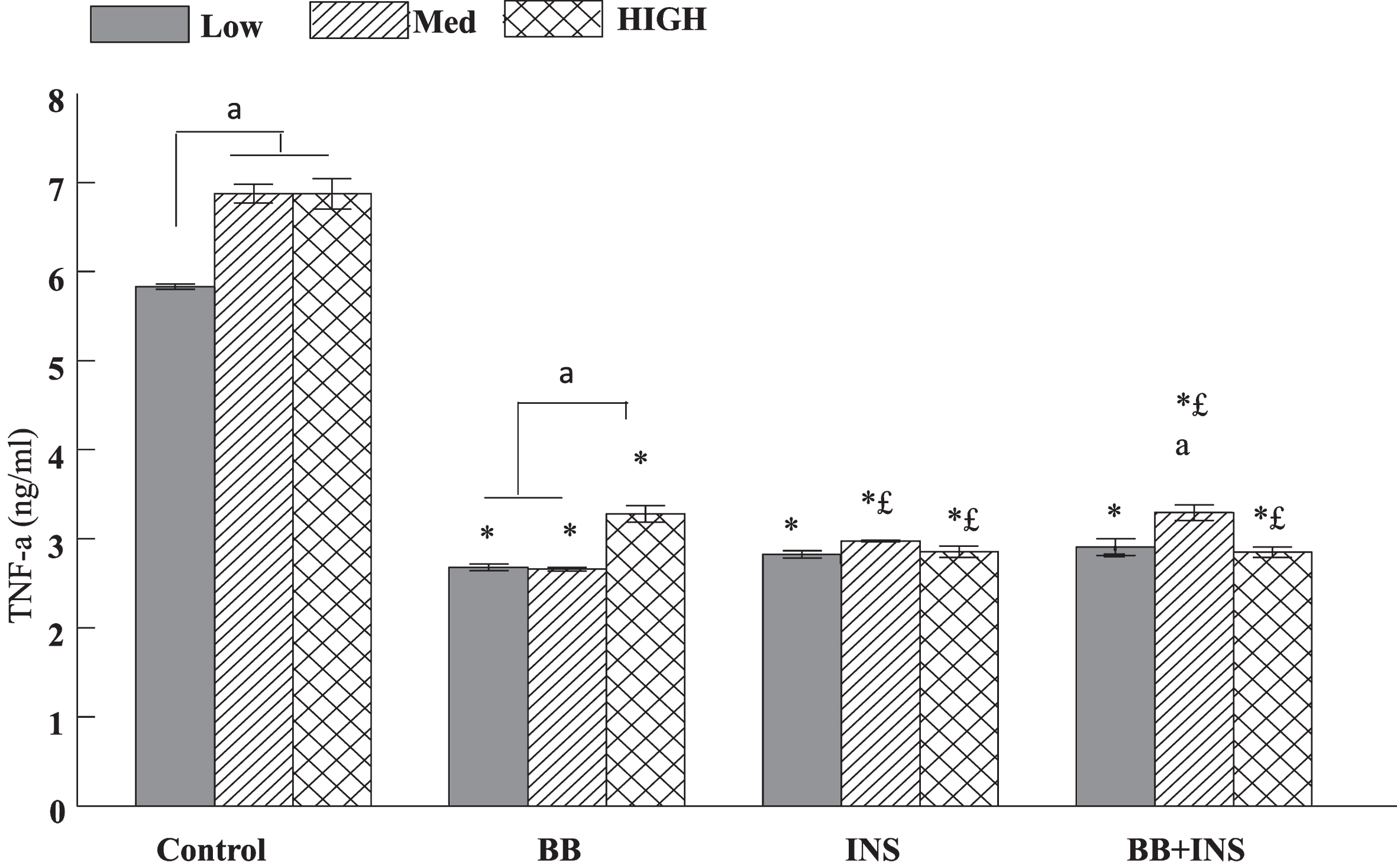
The assessments are presented only with LPS treatment due to the weak production of this cytokine without LPS. LPS-induced levels of TNF-α, a pro-inflammatory cytokine, were significantly increased by increased glucose concentration (P < 0.0001) but reduced by BB (P < 0.0001) and INS (P < 0.0001) treatments (Fig. 3). Post-hoc analysis indicated significant increases in TNF-α release following LPS treatment with medium and high glucose concentrations (Fig. 3) while BB, INS and the combination (INS and BB) significantly reduced TNF-α secretion when compared to glucose-matched LPS control. When the cells were treated with INS alone or the combination of INS and BB, INS was able to significantly reduce TNF-α secretion at the high glucose concentration compared to the glucose-matched BB (P < 0.001), but at the medium glucose concentration, BB alone was the most effective (compared to INS alone or the combination of INS and BB: p < 0.05) (Fig. 3).
Fig. 3
Production of extracellular TNF-α in HAPI microglial cells cultured media containing different concentrations of glucose and treated with 2% blueberries (BB), 50 mM insulin (INS), or combination of blueberries and insulin for 6 hours before LPS treatment. The data were assayed by ELISA and the values are expressed as mean±SEM. Treatment groups were compared with a level of significance at p < 0.05. The symbols mean a statistical difference between treatments. Data sharing the same letters are not different. a means statistical significance between medium or high glucose vs low glucose in each treatment; * means a statistical significance between treatment vs Control (= LPS treatment) respectively for each glucose concentration; £ means a statistical difference between BB vs INS or BB + INS. In control group submitted only to LPS treatment, TNF−α was significantly increased in medium and high glucose medium compared to low glucose (a < 0.05). In BB and Ins and BB + INS pretreated microglia, TNF-α was significantly lower than control at each glucose concentration (*< 0.05). Furthermore, in INS and BB+INS the level of TNF-α was significantly different from BB alone (£< 0.05)
3.4Expression of NOX4
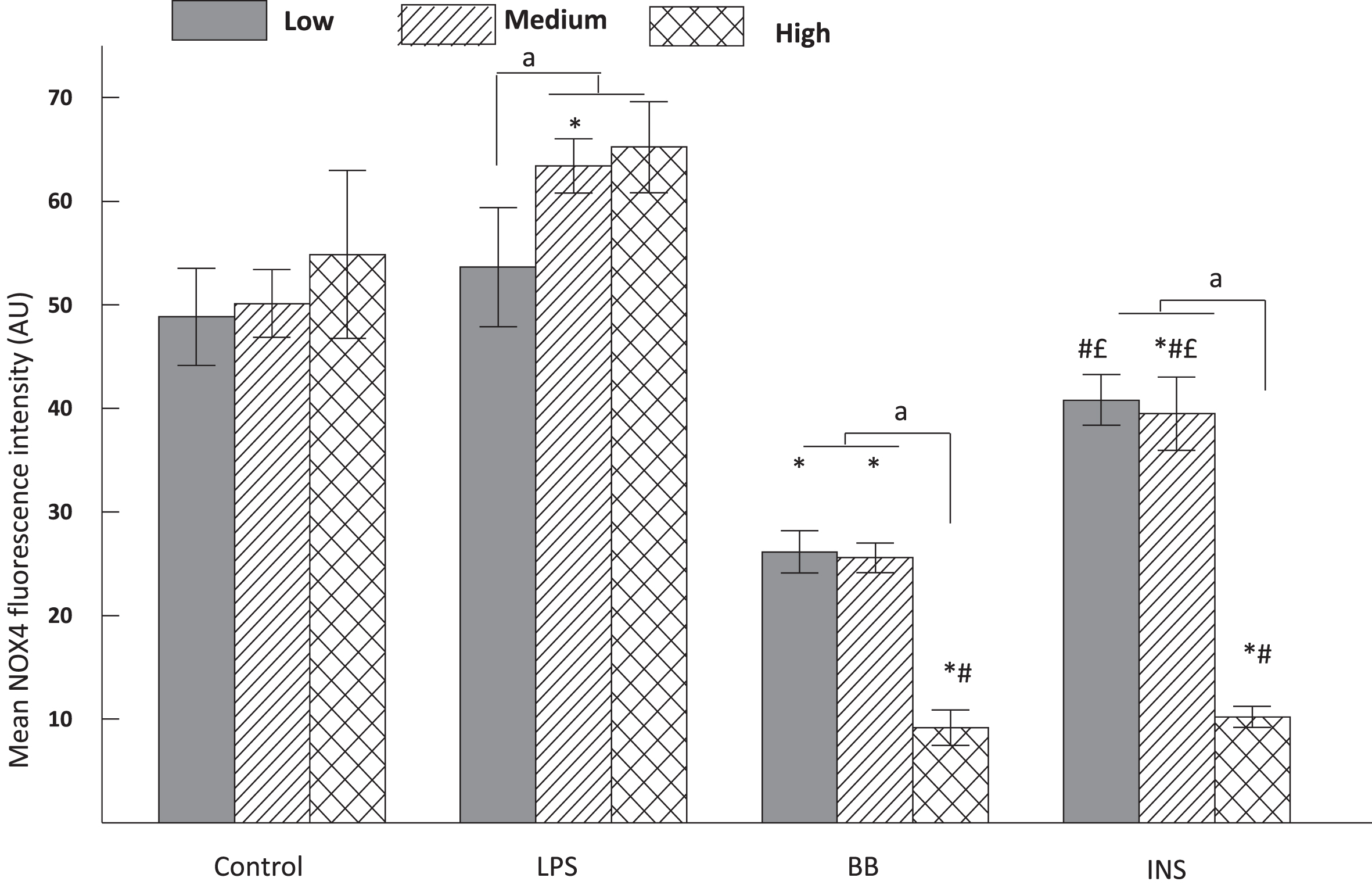
We hypothesized that NADPH oxidase isoform-4 (NOX4), which produces superoxide radical mediating the production of neurotoxic and neurotrophic factors might be involved in the inflammation process associated with glucose. We observed that microglia NOX4 expression was not significantly different between the glucose concentrations but significantly affected by LPS treatment (F1,70 = 6.23, P = 0.01). Post-hoc comparisons indicated that LPS significantly upregulated NOX4 expression under medium and high glucose conditions vs low glucose (P < 0.05). The main effects were significant for BB (F1,60 = 109.16, P < 0.0001) and Post-hoc comparisons indicated that Blueberry treatment significantly attenuated LPS-induced NOX4 upregulation, regardless of glucose concentration (p < 0.05) (Fig. 4). Similarly, insulin also attenuated LPS-induced upregulation of NOX4 (F1,71 = 47.48, P < 0.0001), but the effect of INS had a lesser extent compared to BB (Fig. 4): indeed under low and medium glucose concentrations, the BB treatment showed a significantly greater attenuation of NOX4 expression compared to INS (P < 0.05). The treatment with BB and INS in combination was not performed in this experiment.
Fig. 4
Expression of NOX4 in HAPI cells cultured under different concentrations of glucose and pretreated with control, lipopolysaccharide (LPS), blueberries (BB), 50 mM insulin (INS) for 4 hours. The cultured cells in chamber slides were fixed with methanol and labeled for NOX4 immunofluorescence. The mean intensity of NOX4 from randomly selected 10–15 cells were represented as mean±SEM and compared against LPS (100 ng/mL) treatment alone. The comparative post-hoc analysis was done with significance at p < 0.05. The symbols mean a statistical difference between treatments. Data sharing the same letter or symbols are not different. The letter a means statistical significance between medium or high glucose vs low glucose in each treatment; * means a statistical difference between treatments vs Control respectively for each glucose concentration; # means a statistical difference between treatments vs LPS as a positive control respectively for each glucose concentration; the letter £ means a statistical difference between BB vs others treatments (INS and INS+BB) respectively for each glucose concentration. In control group NOX4 was not significantly different between the glucose concentrations. In LPS stimulated microglia, NOX4 was significantly higher in medium and high glucose (a < 0.05) compared to low glucose. In the BB group, NOX 4 was significantly lower at each glucose condition compared to the control group (b < 0.05) and to LPS (c < 0.05). Ins pretreated cells have a lower NOX level compared to LPS (c < 0.05) in each glucose condition and the level of NOX4 were significantly higher than those in the BB at low and medium glucose (d < 0.05).
3.5Expression of GLUT1
The effect of glucose concentration on GLUT-1 expression was assessed on Hapi-cells stimulated or not with LPS treatment. We observed that with increasing glucose concentration, the GLUT1 decreased (F2,81 = 31.64, P < 0.0001; Fig. 5). Interestingly, treatment with LPS induced up-regulation of GLUT1 as reflected by a significant main effect of LPS (F2,81 = 90.99, P < 0.0001) and Post-hoc treatment indicated that the increase in GLUT1 was significant only in medium and high glucose conditions compared to glucose-matched concentrations as control (Fig. 5). Treatment with BB and INS individually interacted significantly with LPS leading to a significant main effect of BB (P < 0.0001) and INS (P < 0.0001) and Post-hoc analysis indicated that BB and INS treatments were able to significantly reduce GLUT1 at medium glucose concentrations. Additionally, at low glucose concentration, only INS are able to increase GLUT-1. The Fig. 6 presents a part of the immunofluorescence analysis for the three glucose concentrations with and without LPS.
Fig. 5
Expression of glucose trasporter-1 (GLUT1) in HAPI cells cultured in different concentrations of glucose and pretreated with control, lipopolysaccharide (LPS), blueberries (BB), insulin (INS) for 4 hours. The cultured cells in chamber slides were fixed with methanol and labeled for GLUT1 immunofluorescence. The mean intensity of GLUT1 from randomly selected 10–15 cells were represented as mean±SEM and compared against glucose as well as glucose matched treatments with significance p < 0.05. The letter and symbols mean a statistical difference between treatments. The letter a means statistical difference between medium or high glucose vs low glucose for each treatment; * means a statistical difference between treatments vs Control respectively for each glucose concentration; “#” means a statistical difference between treatments vs LPS as a positive control respectively for each glucose concentration; the letter “£” means a statistical difference between BB and INS respectively to glucose concentration. The level of GLUT1 is significantly decreased in medium and high glucose compared to low glucose (a < 0.05) in control and INS. Compared to LPS, there were significantly differences in the level of GLUT1 in BB and INS (c < 0.05) at medium. At low glucose concentration, INS has a significant effect on GLUT-1.
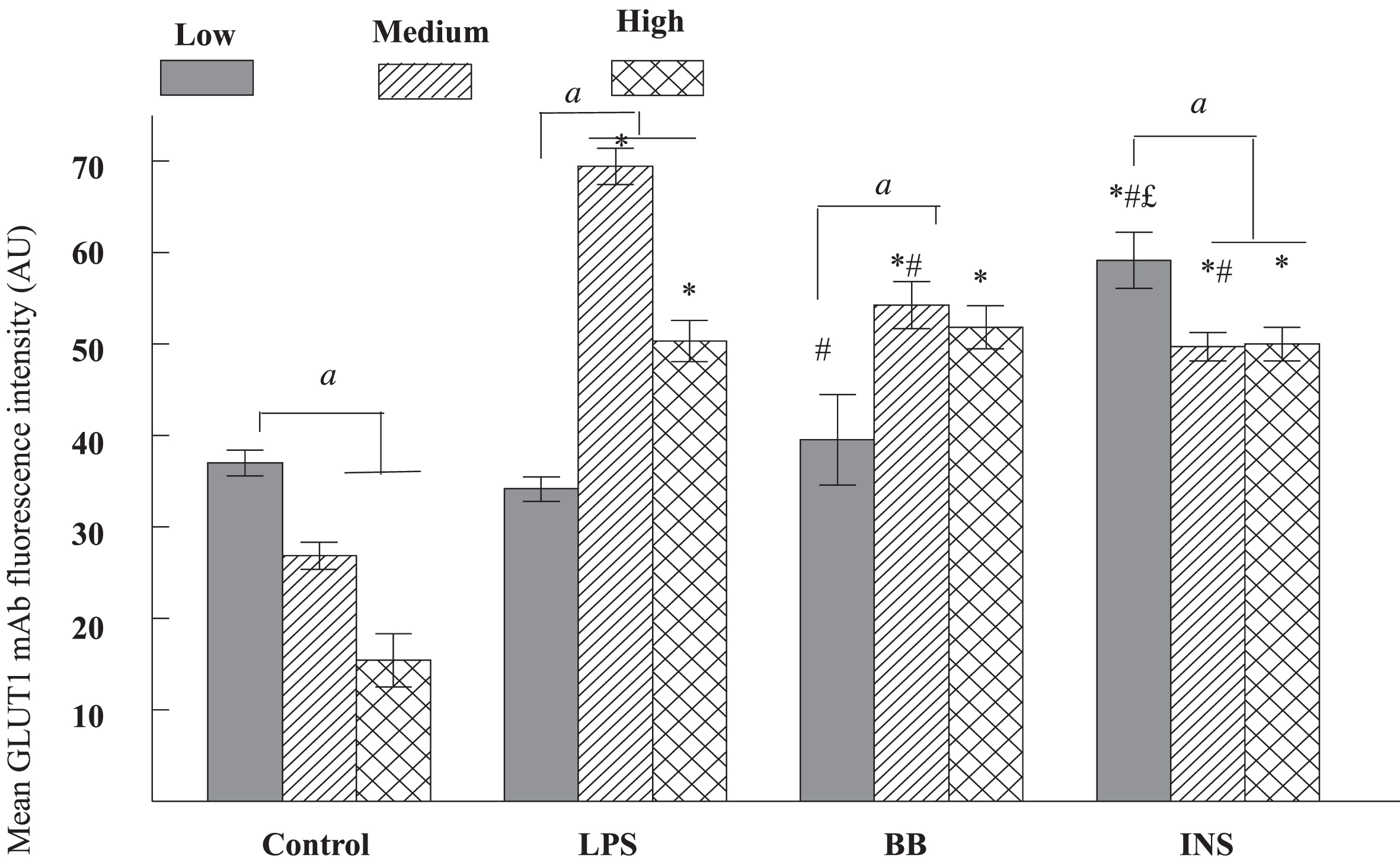
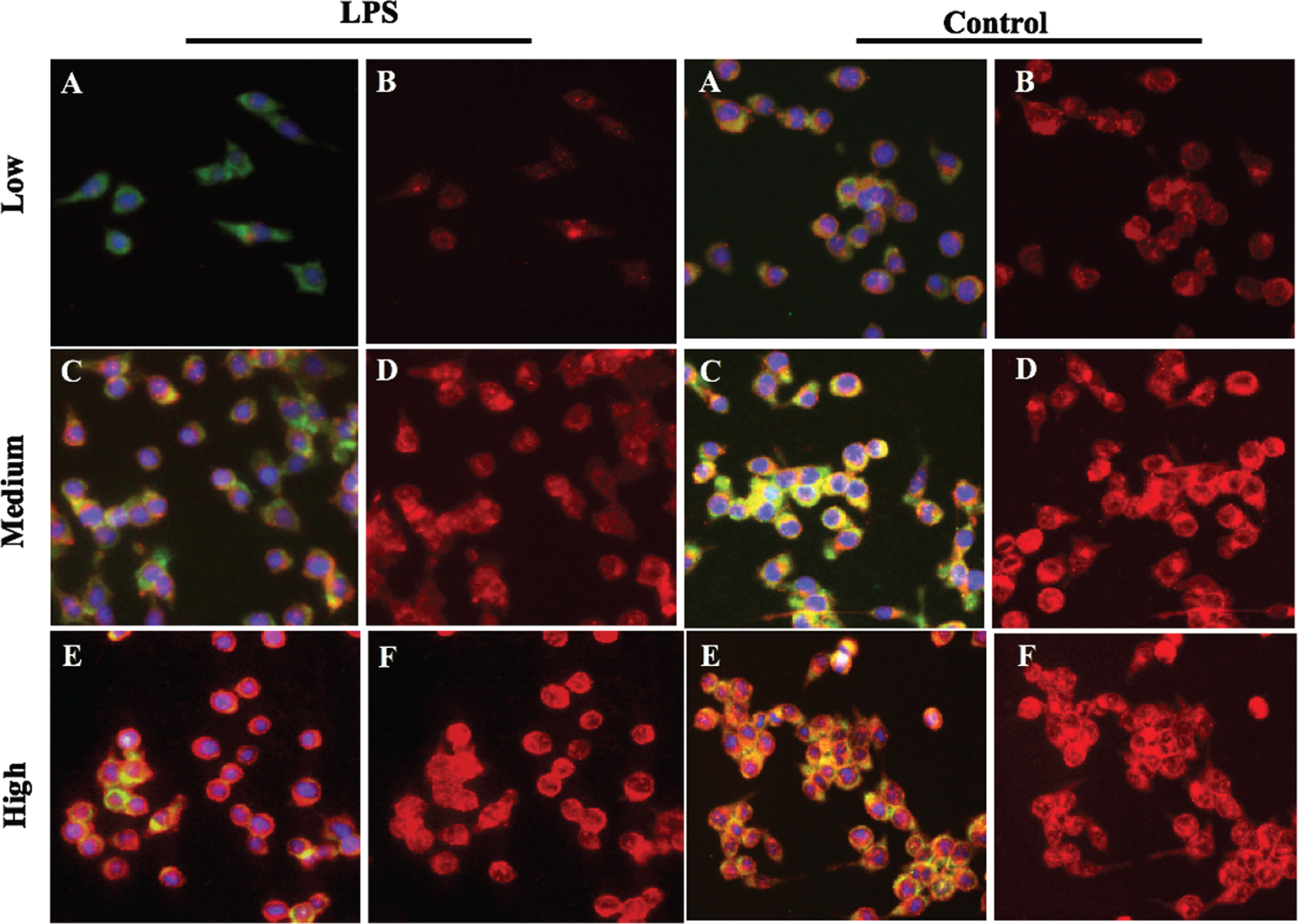
Fig. 6
Immunofluorescence analysis for GLUT-1 expression in HAPI cells. HAPI cells on culture slides were incubated with either low (5 mM), medium (25 mM) or high (50 mM) glucose in Dulbecco’s modified Eagle’s medium, with or without LPS. After 2 hours of LPS treatment the cells were washed with PBS, fixed in 100% cold methanol, incubated overnight at 4°C with GLUT1 antibody (Upstate-Millipore, Billerica, MA), and co-stained with β-actin. Species-specific TRITC or FITC-conjugated secondary antibodies were used to stain GLUT-1 (green: A,C,E) and β-actin (red: B,D,F). DAPI is used for nuclear staining. Supporting the quantitative data in Fig. 5, immunostaining of Glut 1 clearly indicates that GLUT1 expression was significantly lowered with high glucose as well as LPS treatments.
4Discussion
In this study, we observed that increased glucose concentration can exacerbate microglia-mediated neuroinflammation, evidenced by an overproduction of NO and the up-expression of several pro-oxidant and pro-inflammatory mediators such as iNOS, TNF-α and NOX4 that are capable of producing ROS. We also demonstrated that BB and insulin were capable of alleviating these effects induced by increased glucose level, particularly when microglia are challenged by LPS, suggesting that they could potentially improve the neuronal environment and overall brain health. Indeed, targeting microglia is important to control neuroinflammation, since microglia activation is known to be implicated in many neurodegenerative diseases. In fact, the term “neuroinflammation” is in some cases replaced with “microglia activation”, owing to its key role in response to neural dysfunction [36].
4.1Anti-inflammatory effects of BB and Insulin in high glucose conditions
We observed a significant effect of glucose on iNOS overexpression, which was even more pronounced following stimulation with LPS. Our data showed an over-production of NO when microglia were cultured in high glucose conditions and were activated by LPS. Given that the increase in iNOS expression is known to cause tissue damage in the CNS through increased production of NO, which can form peroxynitrites, a harmful ROS implicated in the pathogenesis of many diseases, including neurodegenerative diseases [37], our data suggest a higher risk when inflammation takes place in a cell environment with high glucose. Our results on microglia affirm those of a previous study on pulmonary microvascular endothelial cells which showed that hyperglycemia significantly increased LPS-stimulated iNOS production and enhanced LPS-stimulated nitrite/nitrate production [38]. Over-production of NO was also demonstrated in rat mesangial cells exposed to high glucose via increasing iNOS [39]. Our data with insulin treatment showed reduced inflammation followed by decreased NO, iNOS and TNF-α, suggesting a protective anti-inflammatory effect of insulin against neuroinflammation. Interestingly, in accordance with our results, brain specimens from patients with uncontrolled hyperglycemia who died in intensive care showed a strong increase in microglia activation [40], while preventing hyperglycemia with insulin reduced abnormalities, with microglial activation being the earliest preventable event [41]. Also in agreement with us, a recent study showed reduced NO, ROS, and TNF-α production, as well as iNOS expression, in BV2 microglia stimulated with LPS, followed by insulin administration [42].
Our results indicate that not only insulin but also BB treatment significantly reduced both reactive nitrite production and iNOS expression compared to LPS-only controls, particularly under medium glucose levels. Studies from our laboratory and others have shown that blueberries, which are rich in anthocyanins and numerous other phytonutrients, protect brain cells and improve cognitive functions through reduction of ROS burden and neuroinflammation, while improving protective neuronal signaling [43–47]. The BB treatment at high glucose conditions did not elicit the same or proportional attenuation in LPS-induced nitrite production or iNOS expression as insulin, possibly due to the sugar content in these extracts, which could have overshadowed the antioxidant benefits at these glucose concentrations. We observed no additional or synergistic effect of BB and insulin when both treatments were administered, as if the beneficial pattern was similar to the INS treatment alone. One possibility is that BB constituents and Ins might trigger the signaling pathway to the IGF-1 receptor, given that both are known to modulate these receptors [33]. We previously reported that BB treatment reduced proinflammatory protein mediators via mRNA IGF-1 in microglia [48]. In agreement with these data, BB feeding is reported to upregulate IGF-1 in aged [18] and young rat [49] brains compared to control-fed animals. Furthermore, it has been shown that IGF-1 treatment can mitigate the production of ROS and TNF-α in activated microglia [50]. Therefore, we assumed that BB extract might contain polyphenols that work as insulin-like molecules and might mediate anti-inflammatory activity through the insulin/IGF-1 pathway [51].
The proinflammatory cytokine, TNF-α, is a key mediator in stimulating the production and release of neurotoxic mediators. TNF-α stimulates a rapid accumulation of the neurotoxic lipid messenger, ceramide, and overproduction of ROS, primarily through NOX activities [52]. Under normal and high glucose concentration, LPS significantly increased TNF-α production, which was effectively attenuated by pre-treatments with BB or INS. These findings agree with previous studies where BB reduced LPS-induced TNF-α in BV2 microglial cells [33], and also in mice where BB alleviates inflammation induced by a high fat diet through a decrease in TNF-α [53]. In accordance with Brabazon et al. (2018) [42], who for the first time reported that INS acts on microglia (BV2), and in particular significantly reduces TNF-α release following LPS-stimulation, we observed a similar effect on Hapi cells. Taken together these data validate our cell model for studying the impact of a nutritional strategy to modulate neuro-inflammation and show that BBs might be a good surrogate for INS to reduce TNF-α expression. This blunted effect on these pro-inflammatory cytokines might be a part of the protective effect of BB against NOX.
Members of the NOX family have gained interest as sources of oxidative stress in the aging process, mild cognitive disorders, and numbers of chronic pathologies. While we previously observed that BB is capable of blunting NOX2 expression [44], which was confirmed by others [54], data on NOX4 are rare. In the present study, NOX4 expression was found to be upregulated under higher glucose conditions following LPS stimulation. To our knowledge, this is the first time this upregulation of glucose-induced NOX4 has been reported in microglia, and the first to report that INS and BB can attenuate this increase. In agreement with our data, previous studies showed that NOX4 was upregulated in renal cortex under diabetic conditions, [55] and in retinal endothelial cells under high glucose (30 mmol/L) compared to normal glucose (5.5 mmol/L) [56]. Although in our study this effect was observed only with LPS-stimulation, our results are also in agreement with a previous study from Suh and coll. [57] where neuronal NOX activation was linked to glucose and neuronal death that occurred during ischemia-reperfusion.
We also observed that BBs inhibit NOX4 expression and that this inhibition is greater with high glucose. This result is in accordance with Huang et al. (2018) who reported a decrease in NOX4 expression with BB in retinal endothelial cells grown in high glucose [56]. It has been previously shown that BB could disrupt functional assembly of NOX in lipid rafts of neuronal cells exposed to TNF-α [54], which could be the mechanism by which BBs act to inhibit NOX4 overexpression, even if we cannot exclude an antioxidant effect. Furthermore, we observed that insulin also decreased NOX4 expression induced by high glucose. In accordance with this effect of insulin on microglia, a study has reported that rosiglitazone, an anti-diabetic molecule, reduced NOX4 upregulation in hyperglycemia-activated endothelial cells [58]. In this study the effect was mediated by NF-kB on NOX4; therefore modulation of NF-kB activation may be the common pathway for both BBs and INS to reduce NOX4 expression in microglia under high glucose conditions. Taken as a whole, BB and insulin treatments may reduce the oxidative burden in microglial cells either through disruption of NOX complex formation, by direct quenching of superoxide, or by attenuating NOX4 transcript levels. Considering the high toxicity of NOX4, managing its expression might offer benefits against neuroinflammation.
4.2BB and insulin dampen the glucose effect on GLUT1 expression
GLUT-1 has been identified as playing an important role in intracellular glucose uptake in microglia activation [14, 59]. Deficiency in GLUT-1 is associated with delayed psychomotor development, microcephaly, ataxia, spasticity and movement disorders [60]. Our results indicated that high glucose concentrations down-regulated GLUT-1 expression, while LPS treatment up-regulated GLUT-1 expression. In agreement with our data a study reports that glucose uptake in microglia is facilitated by GLUT-1, particularly under inflammatory conditions [14]. The goal of this effect is to facilitate glucose uptake by microglia and promote anaerobic glycolysis under inflammatory conditions. Previous studies have also demonstrated that the pro-inflammatory cytokine interleukin (IL)-1β upregulates GLUT1 expression in microglia [61] and that up-regulation allowed the cells to meet the increased energy demand during an immunological challenge, by increasing the amount of glucose entering the cells [62]. Interestingly, we observed decrease in GLUT1 expression by BB in a way similar to insulin at medium glucose. Blocking GLUT-1 has been considered as an effective approach to re-program the metabolic pathway and control microglia activation and neurodegeneration [14]. Indeed, it is thought that blocking glucose may not only suppress microglia activation but also increase glucose uptake by GLUT-3 neurons, in the inflamed ischemic CNS when glucose supply is restricted [14]. Thus, BB might play a role only when glucose disorders are present, because the low glucose in our study is a normal glucose level for the brain. This specific effect of BBs might be compared with the reported role of BB in improving insulin resistance in a population with diabetes type 2 or an experimental model of such, but to our knowledge there is no description of BB inducing hypoglycemia in healthy individuals with normal glycemia [16, 19, 22, 30, 64]. To our knowledge, this is the first time that an effect of BB was reported on GLUT1 in microglia. Another study in adipose tissue [63] saw that chokeberry, a fruit with a similar phytochemical profile as that of blueberry, increased insulin signaling and GLUT-1 expression, and decreased inflammatory cytokines.
In the current study, only INS was able to increase GLUT1 under low glucose conditions, in agreement with the well-known role of insulin, which is to normalize glucose concentration in brain. We are unable to explain the lack of effect of both BB and INS on LPS at high glucose concentration.
Taken as a whole, our data propose BB supplementation as a new preventive and therapeutic strategy for brain health and the treatment of neurodegenerative disease associated with high glucose. Obviously, this in vitro preliminary study is limited, and future studies could measure glucose uptake and the effect of BBs on mRNA expression of the INS receptor and TLR4 receptors. The results of this study must be confirmed in other cell models, such as neurons, in order to evaluate the effect of BBs on neurogenesis and BDNF (Brain-Derived Neurotrophic Factor) expression in comparison to INS, to evaluate how BB supplementation could be a good alternative to stimulate brain function.
5Conclusion
This work demonstrates that hyperglycemia enhances microglial ROS production and inflammation responses, which are known to have deleterious effects in the CNS. Therefore, it is essential to elucidate strategies that are safe and effective in mitigating the hyperglycemia-induced neuro-inflammation and ROS overproduction in the brain, especially in the brain of those with metabolic syndrome. For the first time this study compared BBs and INS on their ability to reduce the pro-inflammatory response of microglia. Our data suggest that both insulin and blueberries were effective in reducing ROS and inflammatory burden in microglia, and this may be one mechanism by which BB and insulin have beneficial effects in CNS injury or neurodegenerative conditions. Both treatments were also able to decrease GLUT-1 mRNA expression induced by LPS, even under high glucose conditions. These data suggest that BB polyphenols have effects on GLUT-1, and this may be another mechanism by which BB acts to prevent neurodegenerative decline. Given the increasing prevalence of age-associated disorders in the population, including diabetes type 2 and undiagnosed high blood glucose, BB supplementation to protect the brain from inflammation induced by hyperglycemia may represent an important alternative to insulin as a therapeutic option to preserve brain health.
Although more studies are needed to understand the exact mechanisms involved, exploring dietary approaches such as blueberries to improve brain health during hyperglycemia could prove to be an effective and viable strategy, before patients need insulin therapy.
Acknowledgments
The authors would like to thank the personnel of the Neuroscience and Aging Lab, HNRCA.
Funding
This research was supported by funds from University of Grenoble Alpes, France (Floralis A10CUF00294000).
Conflicts of interest
The authors declare to have no conflict of interest.
References
[1] | Kim B , Feldman EL . Insulin resistance in the nervous system. Trends Endocrinol Metab. (2012) ;23: (3):133–41. |
[2] | Samaras K , Lutgers HL , Kochan NA , Crawford JD , Campbell LV , Wen W , et al. The impact of glucose disorders on cognition and brain volumes in the elderly: the Sydney Memory and Ageing Study. Age (Dordr). (2014) ;36: (2):977–93. |
[3] | Liu Z , Patil IY , Jiang T , Sancheti H , Walsh JP , Stiles BL , et al. High-fat diet induces hepatic insulin resistance and impairment of synaptic plasticity. PLoS One. (2015) ;10: (5):e0128274. |
[4] | Verdile G , Keane KN , Cruzat VF , Medic S , Sabale M , Rowles J , et al. Inflammation and Oxidative Stress: The Molecular Connectivity between Insulin Resistance, Obesity, and Alzheimer’s Disease. Mediators Inflamm. (2015) ;2015: :105828. |
[5] | Molina MN , Ferder L , Manucha W . Emerging Role of Nitric Oxide and Heat Shock Proteins in Insulin Resistance. Curr Hypertens Rep. (2016) ;18: (1):1. |
[6] | Han X , Ma Y , Liu X , Wang L , Qi S , Zhang Q , et al. Changes in insulin-signaling transduction pathway underlie learning/memory deficits in an Alzheimer’s disease rat model. J Neural Transm (Vienna). (2012) ;119: (11):1407–16. |
[7] | Geijselaers SLC , Sep SJS , Stehouwer CDA , Biessels GJ . Glucose regulation, cognition, and brain MRI in type 2 diabetes: a systematic review. Lancet Diabetes Endocrinol. (2015) ;3: (1):75–89. |
[8] | Biessels GJ , Bravenboer B , Gispen WH . Glucose, insulin and the brain: modulation of cognition and synaptic plasticity in health and disease: a preface. Eur J Pharmacol. (2004) ;490: (1-3):1–4. |
[9] | Ghosh S , Castillo E , Frias ES , Swanson RA . Bioenergetic regulation of microglia. Glia. (2018) ;66: (6):1200–12. |
[10] | Liu Y , Li M , Zhang Z , Ye Y , Zhou J . Role of microglia-neuron interactions in diabetic encephalopathy. Ageing Res Rev. (2018) ;42: :28–39. |
[11] | Lull ME , Block ML . Microglial activation and chronic neurodegeneration. Neurotherapeutics. (2010) ;7: (4):354–65. |
[12] | Bordt EA , Polster BM . NADPH oxidase- and mitochondria-derived reactive oxygen species in proinflammatory microglial activation: a bipartisan affair? Free Radic Biol Med. (2014) ;76: :34–46. |
[13] | Sorce S , Krause KH , Jaquet V . Targeting NOX enzymes in the central nervous system: therapeutic opportunities. Cell Mol Life Sci. (2012) ;69: (14):2387–407. |
[14] | Wang L , Pavlou S , Du X , Bhuckory M , Xu H , Chen M . Glucose transporter 1 critically controls microglial activation through facilitating glycolysis. Mol Neurodegener. (2019) ;14: (1):2. |
[15] | Chen Y , Zhao Y , Dai CL , Liang Z , Run X , Iqbal K , et al. Intranasal insulin restores insulin signaling, increases synaptic proteins, and reduces Abeta level and microglia activation in the brains of 3xTg-AD mice. Exp Neurol. (2014) ;261: :610–9. |
[16] | Shishtar E , Rogers GT , Blumberg JB , Au R , Jacques PF . Long-term dietary flavonoid intake and risk of Alzheimer disease and related dementias in the Framingham Offspring Cohort. Am J Clin Nutr. 2020. |
[17] | Kalt W , Cassidy A , Howard LR , Krikorian R , Stull AJ , Tremblay F , et al. Recent Research on the Health Benefits of Blueberries and Their Anthocyanins. Adv Nutr. (2020) ;11: (2):224–36. |
[18] | Casadesus G , Shukitt-Hale B , Stellwagen HM , Zhu X , Lee HG , Smith MA , et al. Modulation of hippocampal plasticity and cognitive behavior by short-term blueberry supplementation in aged rats. Nutr Neurosci. (2004) ;7: (5-6):309–16. |
[19] | Miller MG , Hamilton DA , Joseph JA , Shukitt-Hale B . Dietary blueberry improves cognition among older adults in a randomized, double-blind, placebo-controlled trial. Eur J Nutr. (2018) ;57: (3):1169–80. |
[20] | Gildawie KR , Galli RL , Shukitt-Hale B , Carey AN . Protective Effects of Foods Containing Flavonoids on Age-Related Cognitive Decline. Curr Nutr Rep. (2018) ;7: (2):39–48. |
[21] | Wedick NM , Pan A , Cassidy A , Rimm EB , Sampson L , Rosner B , et al. Dietary flavonoid intakes and risk of type 2 diabetes in US men and women. Am J Clin Nutr. (2012) ;95: (4):925–33. |
[22] | Stote KS , Wilson MM , Hallenbeck D , Thomas K , Rourke JM , Sweeney MI , et al. Effect of Blueberry Consumption on Cardiometabolic Health Parameters in Men with Type 2 Diabetes: An 8-Week, Double-Blind, Randomized, Placebo-Controlled Trial. Curr Dev Nutr. (2020) ;4: (4):nzaa030. |
[23] | Ahmed SM , Luo L , Namani A , Wang XJ , Tang X . Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim Biophys Acta Mol Basis Dis. (2017) ;1863: (2):585–97. |
[24] | Hwang YP , Choi JH , Yun HJ , Han EH , Kim HG , Kim JY , et al. Anthocyanins from purple sweet potato attenuate dimethylnitrosamine-induced liver injury in rats by inducing Nrf2-mediated antioxidant enzymes and reducing COX-2 and iNOS expression. Food Chem Toxicol. (2011) ;49: (1):93–9. |
[25] | Yahfoufi N , Alsadi N , Jambi M , Matar C . The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients. (2018) ;10: (11). |
[26] | Joseph SV , Edirisinghe I , Burton-Freeman BM . Berries: anti-inflammatory effects in humans. J Agric Food Chem. (2014) ;62: (18):3886–903. |
[27] | Xiong J , Matta FV , Grace M , Lila MA , Ward NI , Felipe-Sotelo M , et al. Phenolic content, anti-inflammatory properties, and dermal wound repair properties of industrially processed and non-processed acai from the Brazilian Amazon. Food Funct. (2020) ;11: (6):4903–14. |
[28] | Forbes-Hernandez TY , Giampieri F , Gasparrini M , Afrin S , Mazzoni L , Cordero MD , et al. Lipid Accumulation in HepG2 Cells Is Attenuated by Strawberry Extract through AMPK Activation. Nutrients. (2017) ;9: (6). |
[29] | Belwal T , Nabavi SF , Nabavi SM , Habtemariam S . Dietary Anthocyanins and Insulin Resistance: When Food Becomes a Medicine. Nutrients. (2017) ;9: (10). |
[30] | Rozanska D , Regulska-Ilow B . The significance of anthocyanins in the prevention and treatment of type 2 diabetes. Adv Clin Exp Med. (2018) ;27: (1):135–42. |
[31] | Lau FC , Shukitt-Hale B , Joseph JA . Nutritional intervention in brain aging: reducing the effects of inflammation and oxidative stress. Subcell Biochem. (2007) ;42: :299–318. |
[32] | Routh VH , Hao L , Santiago AM , Sheng Z , Zhou C . Hypothalamic glucose sensing: making ends meet. Front Syst Neurosci. (2014) ;8: :236. |
[33] | Lau FC , Bielinski DF , Joseph JA . Inhibitory effects of blueberry extract on the production of inflammatory mediators in lipopolysaccharide-activated BV2 microglia. J Neurosci Res. (2007) ;85: (5):1010–7. |
[34] | Poulose SM , Fisher DR , Larson J , Bielinski DF , Rimando AM , Carey AN , et al. Anthocyanin-rich acai (Euterpe oleracea Mart.) fruit pulp fractions attenuate inflammatory stress signaling in mouse brain BV-2 microglial cells. J Agric Food Chem. (2012) ;60: (4):1084–93. |
[35] | Poulose SM , Fisher DR , Bielinski DF , Gomes SM , Rimando AM , Schauss AG , et al. Restoration of stressor-induced calcium dysregulation and autophagy inhibition by polyphenol-rich acai (Euterpe spp.) fruit pulp extracts in rodent brain cells in vitro. Nutrition. (2014) ;30: (7-8):853–62. |
[36] | Graeber MB , Li W , Rodriguez ML . Role of microglia in CNS inflammation. FEBS Lett. (2011) ;585: (23):3798–805. |
[37] | Prabhakar SS . Role of nitric oxide in diabetic nephropathy. Semin Nephrol. (2004) ;24: (4):333–44. |
[38] | Liu XJ , Zhang ZD , Ma XC . High glucose enhances LPS-stimulated human PMVEC hyperpermeability via the NO pathway. Exp Ther Med. (2013) ;6: (2):361–7. |
[39] | Zhai YP , Lu Q , Liu YW , Cheng Q , Wei YQ , Zhang F , et al. Over-production of nitric oxide by oxidative stress-induced activation of the TGF-beta1/PI3K/Akt pathway in mesangial cells cultured in high glucose. Acta Pharmacol Sin. (2013) ;34: (4):507–14. |
[40] | Sonneville R , den Hertog HM , Guiza F , Gunst J , Derese I , Wouters PJ , et al. Impact of hyperglycemia on neuropathological alterations during critical illness. J Clin Endocrinol Metab. (2012) ;97: (6):2113–23. |
[41] | Sonneville R , Vanhorebeek I , den Hertog HM , Chretien F , Annane D , Sharshar T , et al. Critical illness-induced dysglycemia and the brain. Intensive Care Med. (2015) ;41: (2):192–202. |
[42] | Brabazon F , Bermudez S , Shaughness M , Khayrullina G , Byrnes KR . The effects of insulin on the inflammatory activity of BV2 microglia. PLoS One. (2018) ;13: (8):e0201878. |
[43] | Willis LM , Freeman L , Bickford PC , Quintero EM , Umphlet CD , Moore AB , et al. Blueberry supplementation attenuates microglial activation in hippocampal intraocular grafts to aged hosts. Glia. (2010) ;58: (6):679–90. |
[44] | Poulose SM , Bielinski DF , Carrihill-Knoll KL , Rabin BM , Shukitt-Hale B . Protective effects of blueberry- and strawberry diets on neuronal stress following exposure to (56)Fe particles. Brain Res. (2014) ;1593: :9–18. |
[45] | Elks CM , Reed SD , Mariappan N , Shukitt-Hale B , Joseph JA , Ingram DK , et al. A blueberry-enriched diet attenuates nephropathy in a rat model of hypertension via reduction in oxidative stress. PLoS One. (2011) ;6: (9):e24028. |
[46] | Carey AN , Gomes SM , Shukitt-Hale B . Blueberry supplementation improves memory in middle-aged mice fed a high-fat diet. J Agric Food Chem. (2014) ;62: (18):3972–8. |
[47] | Brewer GJ , Torricelli JR , Lindsey AL , Kunz EZ , Neuman A , Fisher DR , et al. Age-related toxicity of amyloid-beta associated with increased pERK and pCREB in primary hippocampal neurons: reversal by blueberry extract. J Nutr Biochem. (2010) ;21: (10):991–8. |
[48] | Shukitt-Hale B , Bielinski DF , Lau FC , Willis LM , Carey AN , Joseph JA . The beneficial effects of berries on cognition, motor behaviour and neuronal function in ageing. Br J Nutr. (2015) ;114: (10):1542–9. |
[49] | Rendeiro C , Vauzour D , Kean RJ , Butler LT , Rattray M , Spencer JP , et al. Blueberry supplementation induces spatial memory improvements and region-specific regulation of hippocampal BDNF mRNA expression in young rats. Psychopharmacology (Berl). (2012) ;223: (3):319–30. |
[50] | Grinberg YY , Dibbern ME , Levasseur VA , Kraig RP . Insulin-like growth factor-1 abrogates microglial oxidative stress and TNF-alpha responses to spreading depression. J Neurochem. (2013) ;126: (5):662–72. |
[51] | Mainardi M , Fusco S , Grassi C . Modulation of hippocampal neural plasticity by glucose-related signaling. Neural Plast. (2015) ;2015: :657928. |
[52] | Barth BM , Stewart-Smeets S , Kuhn TB . Proinflammatory cytokines provoke oxidative damage to actin in neuronal cells mediated by Rac1 and NADPH oxidase. Mol Cell Neurosci. (2009) ;41: (2):274–85. |
[53] | Mykkanen OT , Huotari A , Herzig KH , Dunlop TW , Mykkanen H , Kirjavainen PV . Wild blueberries (Vaccinium myrtillus) alleviate inflammation and hypertension associated with developing obesity in mice fed with a high-fat diet. PLoS One. (2014) ;9: (12):e114790. |
[54] | Gustafson SJ , Dunlap KL , McGill CM , Kuhn TB . A nonpolar blueberry fraction blunts NADPH oxidase activation in neuronal cells exposed to tumor necrosis factor-alpha. Oxid Med Cell Longev. (2012) ;2012: :768101. |
[55] | Sedeek M , Callera G , Montezano A , Gutsol A , Heitz F , Szyndralewiez C , et al. Critical role of Nox4-based NADPH oxidase in glucose-induced oxidative stress in the kidney: implications in type 2 diabetic nephropathy. Am J Physiol Renal Physiol. (2010) ;299: (6):F1348–58. |
[56] | Huang W , Yan Z , Li D , Ma Y , Zhou J , Sui Z . Antioxidant and Anti-Inflammatory Effects of Blueberry Anthocyanins on High Glucose-Induced Human Retinal Capillary Endothelial Cells. Oxid Med Cell Longev. (2018) ;2018: :1862462. |
[57] | Suh SW , Shin BS , Ma H , Van Hoecke M , Brennan AM , Yenari MA , et al. Glucose and NADPH oxidase drive neuronal superoxide formation in stroke. Ann Neurol. (2008) ;64: (6):654–63. |
[58] | Williams CR , Lu X , Sutliff RL , Hart CM . Rosiglitazone attenuates NF-kappaB-mediated Nox4 upregulation in hyperglycemia-activated endothelial cells. Am J Physiol Cell Physiol. (2012) ;303: (2):C213–23. |
[59] | Nijland PG , Michailidou I , Witte ME , Mizee MR , van der Pol SM , van Het Hof B , et al. Cellular distribution of glucose and monocarboxylate transporters in human brain white matter and multiple sclerosis lesions. Glia. (2014) ;62: (7):1125–41. |
[60] | Leen WG , Wevers RA , Kamsteeg EJ , Scheffer H , Verbeek MM , Willemsen MA . Cerebrospinal fluid analysis in the workup of GLUT1 deficiency syndrome: a systematic review. JAMA Neurol. (2013) ;70: (11):1440–4. |
[61] | Jurcovicova J . Glucose transport in brain - effect of inflammation. Endocr Regul. (2014) ;48: (1):35–48. |
[62] | Spolarics Z , Pekala PH , Bagby GJ , Spitzer JJ . Brief endotoxemia markedly increases expression of GLUT1 glucose transporter in Kupffer, hepatic endothelial and parenchymal cells. Biochem Biophys Res Commun. (1993) ;193: (3):1211–5. |
[63] | Qin B , Anderson RA . An extract of chokeberry attenuates weight gain and modulates insulin, adipogenic and inflammatory signalling pathways in epididymal adipose tissue of rats fed a fructose-rich diet. Br J Nutr. (2012) ;108: (4):581–7. |
[64] | Overall J , Bonney SA , Wilson M , Beermann A , Grace MH , Esposito D , et al. Metabolic Effects of Berries with Structurally Diverse Anthocyanins. Int J Mol Sci. (2017) ;18: (2). |


