
The effect of strawberry cultivars, infested with Tetranychus urticae (Acari: Tetranychidae), on the olfactory response of the predatory mite Neoseiulus californicus (Acari: Phytoseiidae)
Abstract
Tetranychus urticae Koch (Acari: Tetranychidae) is a key pest of agricultural crop. One of the best candidates for biological control of this mite is Neoseiulus californicus (McGregor) (Acari: Phytoseiidae). In this study, the olfactory response of the predatory mite to seven strawberry cultivars (‘Marak’, ‘Yalova’, ‘Aliso’, ‘Gaviota’, ‘Sequoia’, ‘Camarosa’ and ‘Chandler’) was studied using a Y-tube olfactometer. We used two-day-old female predatory mites that were reared on spider mite-infested strawberry. The behavioral response of the predator was studied in four experiments: 1) clean air vs. uninfected strawberry plant, 2) clean air vs. spider mite-infested strawberry plant, 3) uninfected strawberry plant vs. spider mite-infested strawberry plant and 4) two-cultivar choice test (spider mite-infested). In the first experiment, the attraction of the predatory mite to uninfected strawberry plant volatiles was significant. In the second test, the predators significantly preferred volatiles from strawberry plant infested with T. urticae to clean air. In the third test, 68% to 80% of the predatory mites were attracted to spider mite-infested strawberry plant volatiles compared with uninfected plants. Among the seven strawberry cultivars, the predator responses to volatiles of infested plants varied 62% to 83% and to volatiles of uninfested plant varied 59% to 75%. Herbivore – induced plant volatile emitted from strawberry infested with T. urticae attract the predatory mite. The last test showed that ‘Aliso’ and ‘Chandler’ were more attractive for the predatory mite.
1Introduction
Plants can respond to herbivore with the production of volatiles (herbivore-induced plant volatiles (HIPV)) that attract carnivorous natural enemies of the herbivorous arthropods [1], this so-called indirect defense affects of plant defense against herbivorous arthropods. Indirect defense affects animal behavior. The plant induced volatiles constitute information that is used in foraging decisions by carnivorous arthropods when searching for herbivorous prey [2]. Behavioral responses of carnivorous arthropods to herbivore induced plant volatiles are flexible and the composition of the blend of volatiles can have a great impact [3, 4]. The olfactory responses of phytoseiid mites are affected by plant species [2, 5], plant developmental stage [6–8], plant cultivar [2, 9, 10], herbivore species [11–13], prey density [8], feeding history [15], predator species [16], odour experience of the predator [14, 15, 17–19] and abiotic factors [6, 18].
Tetranychus urticae is a key pest of agricultural crops [20]. It is a polyphagous herbivore that induces the production of various volatile compounds in different host plants [5]. One of the best candidates for biological control of this mite is Neoseiulus californicus (McGregor) (Acari: Phytoseiidae) [21–24]. The Predatory mite is attracted to spider mite-induced compounds of several plant species such as lima bean [2], apple [10], cucumber [6], tomato [25] and gerbera [26]. The predatory mite is attracted by the herbivore- induced volatiles being released from strawberry plant- infested with Cyclamen mite (Phytonemus pallidus Banks) or leaf beetle Galeracella tenella (L.) over intact plant [27]. In this study, the response of the predatory mite to volatiles associated with different strawberry cultivars infested with T. urticae was examined. The objective of this study was to determine the effect of strawberry volatiles on the foraging behavior of N. californicus in the laboratory. We used a Y-tube olfactometer to test whether odours play a role in prey finding by N. californicus.
22. Materials & methods
Kidney bean, Phaseolus vulgaris L. (‘Alamot’) was used as a food source for T. urticae. The bean plants were grown in a mixture of soil and perlite and were fertilized with N.P.K (20-20-20) every two weeks. Master fertilizer was periodically added. The two-spotted spider mites were collected from a strawberry greenhouse in Karaj (Alborz, Iran). The mites were maintained on kidney bean plants in a climate room (27±1°C, 40±5% RH and 16L: 8D) for more than 2 years.
Seven strawberry cultivars (‘Marak’, ‘Yalova’, ‘Aliso’, ‘Gaviota’, ‘Sequoia’, ‘Camarosa’ and ‘Chandler’) were obtained from University of Tehran (Alborz, Karaj, Iran). Crowns were stored at 1–4°C for 3 weeks, followed by transferring into pots which had a mixture of peat and perlite. The pots were watered daily and fertilized (N-P-K 20:20:20) every two weeks once.
The predatory mite, N. californicus, was obtained from ‘Koppert Biological Systems’ (Berkel en Rodenrijs, The Netherlands) and maintained on leaves of kidney bean that were infested with T. urticae. The stock culture of N. californicus was maintained in a growth chamber (27±1°C, 70±10 R.H. and 16:8 hours L: D). In order to obtain suitable predatory mites for the experiments, the predatory mites were reared on detached leaves of the above-mentioned strawberry cultivars. The leaves were placed upside down on a plastic sheet that was on a water-saturated sponge. Napkin tapes surrounded the plastic sheet, which was put into the water from the other side so that the predatory mites could drink water but could not escape.
To obtain the predatory mites for the experiment, a female predator was taken from the stock colony on lima bean plants and was transferred to spider mite-infested strawberry leaves in a Petri dish. Leaves with fresh prey were replaced every two weeks. Females were allowed to oviposit for one day. Strawberry leaves with fresh prey were replaced every two days. Seven days later, the offspring female was collected. Predators were collected one hour before each bioassay and were kept individually in a Petri dish provided with a droplet of water until the experiment began.
A Y-tube olfactometer [28] was used to test the olfactory response of N. californicus. The olfactometer was made from glass with a Y-shaped metal wire in the middle [29]. The arms were connected via a plastic tube to two Plexiglas boxes (30×30×30 cm) containing odour sources. The air speed inside the arms of the olfactometer was 0.3 m/s that was measured with a digital flow meter. The air containing odour was released separately to each arm of the olfactometer. Each Plexiglas box contained 4 plants of each of the strawberry cultivars, which were carefully removed from the pot soil. The root was gently rinsed with tap water and wrapped in aluminum foil. The female predatory mites were starved for 24 hours, followed by introducing into the base of the olfactometer through a metal wire running through the center of the tube. The behavior of an individual predator was observed for a maximum of 5 min. If the predatory mite moved into one of the arms up to 15 cm, it was considered as a positive response. If it did not reach to the mentioned distance within 5 min, the experiment was stopped. In order to rectify the possible effect of odour location, the odour container side was changed every 7 replicates. For each replication, a fresh predator was used.
Four experiments were conducted using different types of odour sources: A: uninfested plants vs. clean air; B: T. urticae-infested plants vs. clean air; C: T. urticae-infested plants vs. uninfested plants; and D: T. urticae-infested strawberry cultivars were compared with each other. Uninfested strawberry cultivars (without any mite damage) were chosen as ‘control’. To prepare T. urticae-infested strawberry plants, kidney bean leaves (containing ca. 100 T. urticae) were placed on strawberry leaves which were selected for olfactory tests. The plants with spider mite were placed in a climate room (27±1°C, 70±5 R.H. and 16L: 8D) for two days. Each treatment had three replications. For each replicate, 15 predators were used.
The choice data were subjected to a binomial test in Excel. It is deleted the predators that did not choice in olfactometer from the statistical analysis. Statistical analysis was done using a replicated G test which includes a test for heterogeneity among replicates of the experiments [30]. The predators were considered to show preference for either odour source when Gp value were significant,whereas the Gh value was not significant.
3Results
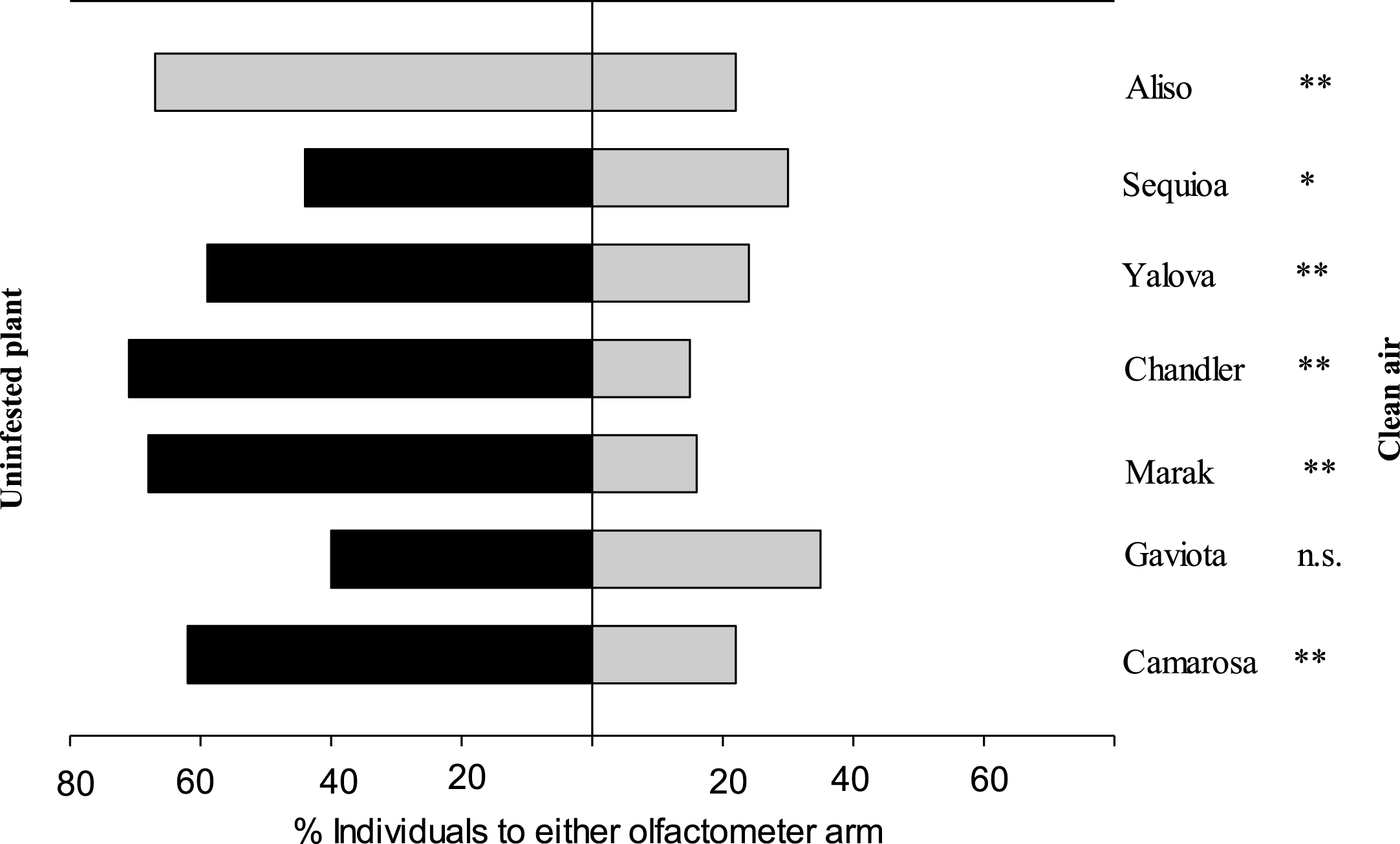
When presented with a choice between unifested strawberry plants (seven cultivars) and clean air. The predator discriminated between these odour source from six cultivars (cultivars: ‘Gaviota’, ‘Camarosa’, ‘Marak’, ‘Chandler’, ‘Aliso’ and ‘Yalova’) (P < 0.05, two binomial test) except for cultivar ‘Sequia’ the predatory mite did not discriminate between the odours (P = 0.18, two binomial test) (Table 1 and Fig. 1).
Table 1
Response of Neoseiulus californicus to odours of uninfested strawberry plants of seven different cultivars vs. clean air in a laboratory two-choice test
| Source of odour | Gh | df | Pvalue | Gp | df | Pvalue | GT | df | Pvalue |
| Uninfested ‘Gaviota’ vs. clean air | 0.90 | 2 | 0.64 | 115.38 | 1 | 0.053* | 2.64 | 3 | 0.20 |
| Uninfested ‘Camarosa’ vs. clean air | 1.35 | 2 | 0.51 | 13.48 | 1 | 0.0002** | 14.53 | 3 | 0.002 |
| Uninfested ‘Marak’ vs. clean air | 0.45 | 2 | 0.80 | 5.75 | 1 | 0.01** | 6.20 | 3 | 0.10 |
| Uninfested ‘Chandler’ vs. clean air | 0.75 | 2 | 0.69 | 7.07 | 1 | 0.008** | 7.82 | 3 | 0.05 |
| Uninfested ‘Aliso’ vs. clean air | 0.03 | 2 | 0.98 | 17.47 | 1 | 0.0002** | 17.50 | 3 | 0.0005 |
| Uninfested ‘Ceqoiua’ vs. clean air | 0.18 | 2 | 0.91 | 1.80 | 1 | 0.18n . s . | 0.18 | 3 | 0.98 |
| Uninfested ‘Yalova’ vs. clean air | 4.17 | 2 | 0.09 | 1.90 | 1 | 0.002** | 6.61 | 3 | 0.08 |
*: P < 0.05, **: P < 0.01, n . s .: not significant.
Fig.1
Responses of adult female of Neoseiulus californicus in Y-tube olfactometer when offer uninfested seven strawberry cultivars leaves v.s. clean air. G test was used to evaluate whether the result differed from a 50 : 50 distribution between the two olfactometer arms (*P < 0.05, **P < 0.01, n.s. not significant).
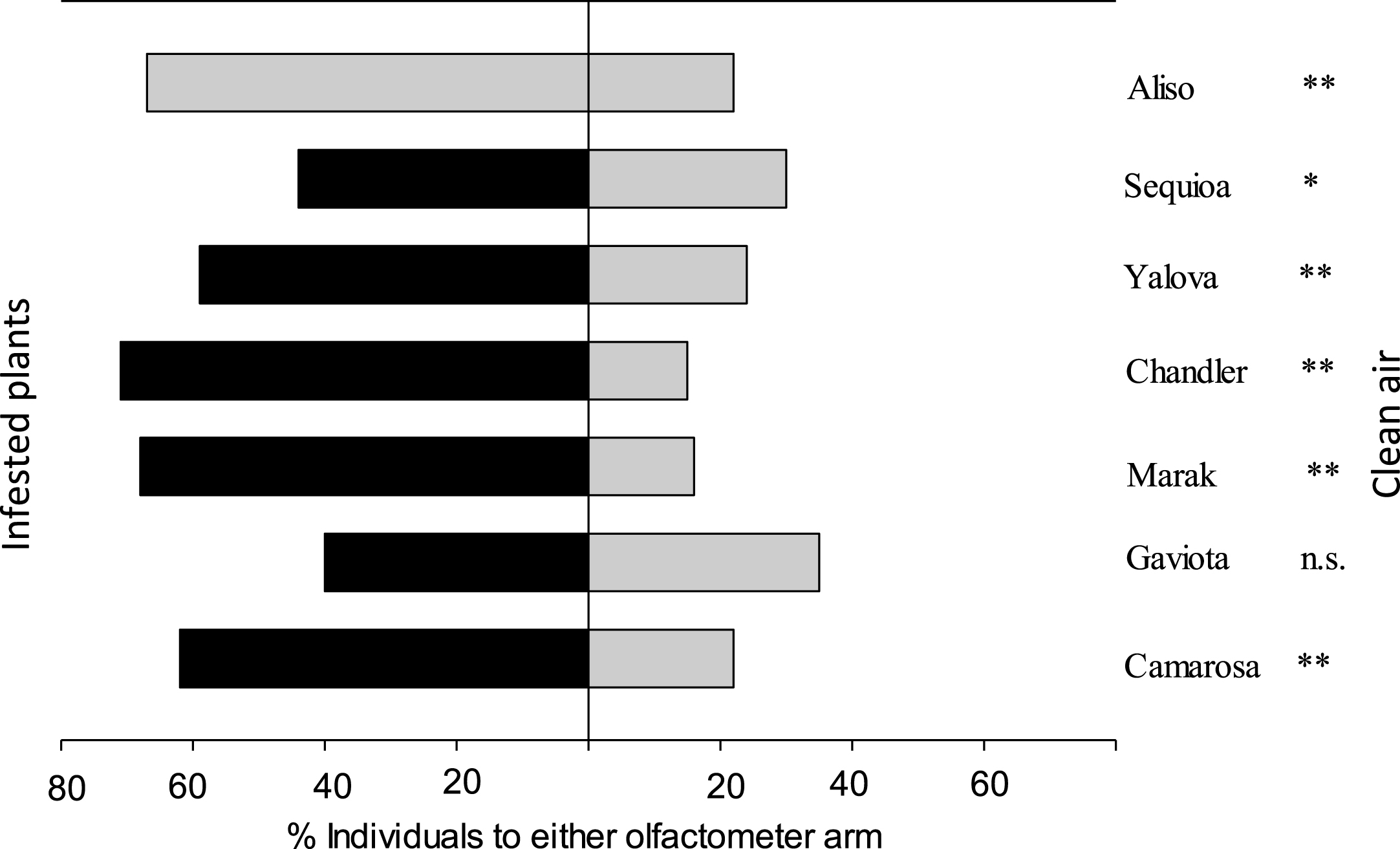
The predator significantly preferred spider mite-infested plants over clean air (P < 0.05, two binomial test) except for ‘Gaviota’ (P = 0.73, two binomial test) (Table 2 and Fig. 2). The percentage of predators preferring the volatiles of infested plants were 73%, 53%, 81%, 82%, 71%, 59% and 75% on ‘Camarosa’, ‘Gaviota’, ‘Marak’, ‘Chandler’, ‘Yalova’, ‘Sequia’ and ‘Aliso’, respectively.
Table 2
Response of Neoseiulus californicus to odours of strawberry cultivars infested with Tetranychus urticae vs. clean air in a laboratory two-choice test
| Source of odour | Gh | df | Pvalue | Gp | df | Pvalue | GT | df | Pvalue |
| ‘Gaviota’ with T. urticae vs. clean air | 1.14 | 2 | 0.57 | 0.18 | 1 | 0.73n . s . | 1.25 | 3 | 0.74 |
| ‘Camarosa’ with T. urticae vs. clean air | 0.21 | 2 | 0.90 | 8.88 | 1 | 0.003** | 9.09 | 3 | 0.03 |
| ‘Marak’ with T. urticae vs. clean air | 2.15 | 2 | 0.34 | 15.39 | 1 | 0.008** | 17.55 | 3 | 0.005 |
| ‘Chandler’ with T. urticae vs. clean air | 1.96 | 2 | 0.37 | 17.36 | 1 | 0.003** | 19.32 | 3 | 0.002 |
| ‘Aliso’ with T. urticae vs. clean air | 1.53 | 2 | 0.46 | 10.46 | 1 | 0.001** | 11.99 | 3 | 0.007 |
| ‘Ceqoiua’ with T. urticae vs. clean air | 0.72 | 2 | 0.70 | 1.13 | 1 | 0.02* | 1.85 | 3 | 0.60 |
| ‘Yalova’ with T. urticae vs. clean air | 0.80 | 2 | 0.67 | 5.94 | 1 | 0.01** | 6.74 | 3 | 0.08 |
*: P < 0.05, **: P < 0.01, n . s .: not significant.
Fig.2
Responses of adult female of Neoseiulus californicus in Y-tube olfactometer when offer infested seven strawberry cultivars leaves v.s. clean air. G test was used to evaluate whether the result differed from a 50:50 distribution between the two olfactometer arms (*P < 0.05, **P < 0.01, n.s. not significant).
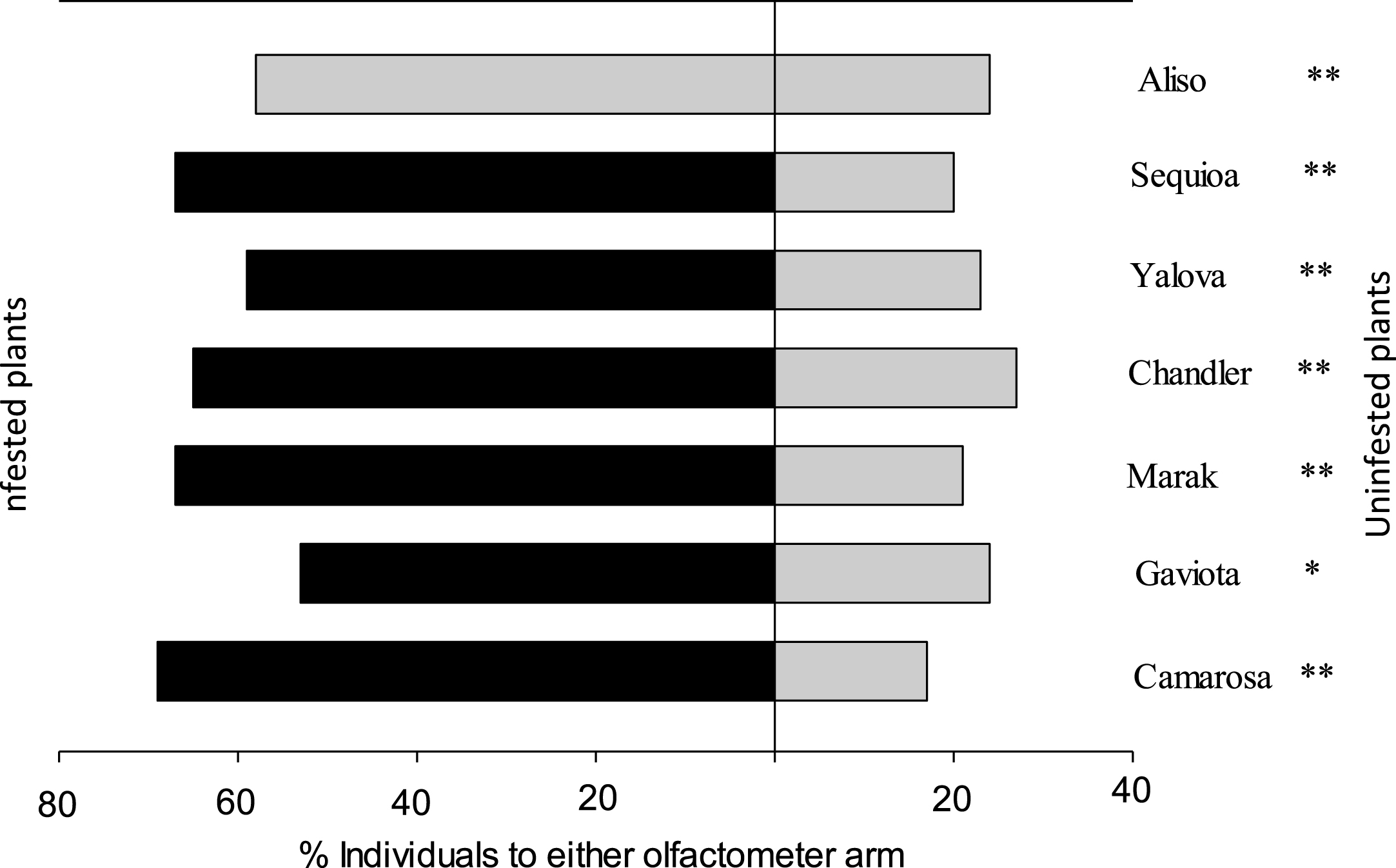
The predator N. californicus significantly preferred odours emitted from plants infested with T. urticae over uninfested plants: 80% on ‘Camarosa’, 68% on ‘Gaviota’, 76% on ‘Marak’, 71% on ‘Chandler’, 73% on ‘Yalova’, 77% on ‘Seqouia’ and 70% on ‘Aliso’ (P < 0.05, two binomial test) (Table 3 and Fig. 3).
Table 3
Response of Neoseiulus californicus to odours of strawberry cultivars infested with Tetranychus urticae vs. uninfested strawberry plant in a laboratory two-choice test
| Source of odour | Gh | df | Pvalue | Gp | df | Pvalue | GT | df | Pvalue |
| ‘Gaviota’ with T. urticae vs. uninfested ‘Gaviota’ | 1.22 | 2 | 0.54 | 4.94 | 1 | 0.03* | 6.16 | 3 | 0.10 |
| ‘Camarosa’ with T. urticae vs. uninfested ‘Camarosa’ | 2.03 | 2 | 0.36 | 14.44 | 1 | 0.0001** | 16.76 | 3 | 0.0009 |
| ‘Marak’ with T. urticae vs. uninfested ‘Marak’ | 0.70 | 2 | 0.70 | 10.24 | 1 | 0.001** | 10.34 | 3 | 0.012 |
| ‘Chandler’ with T. urticae vs. uninfested ‘Chandler’ | 0.65 | 2 | 0.72 | 7.27 | 1 | 0.007** | 7.92 | 3 | 0.05 |
| ‘Aliso’ with T. urticae vs. uninfested ‘Aliso’ | 1.26 | 2 | 0.53 | 6.26 | 1 | 0.012** | 7.52 | 3 | 0.06 |
| ‘Ceqoiua’ with T. urticae vs. uninfested ‘Ceqoia’ | 0.43 | 2 | 0.81 | 11.93 | 1 | 0.0005** | 12.36 | 3 | 0.006 |
| ‘Yalova’ with T. urticae vs. uninfested ‘Yalova’ | 1.93 | 2 | 0.38 | 7.36 | 1 | 0.007** | 9.30 | 3 | 0.03 |
*: P < 0.05, **: P < 0.01, n . s .: not significant.
Fig.3
Responses of adult female of Neoseiulus californicus in Y-tube olfactometer when offer infested seven strawberry cultivars leaves v.s. uninfested seven strawberry cultivars leaves. G test was used to evaluate whether the result differed from a 50 : 50 distribution between the two olfactometer arms (*P < 0.05, **P < 0.01, n.s. not significant).
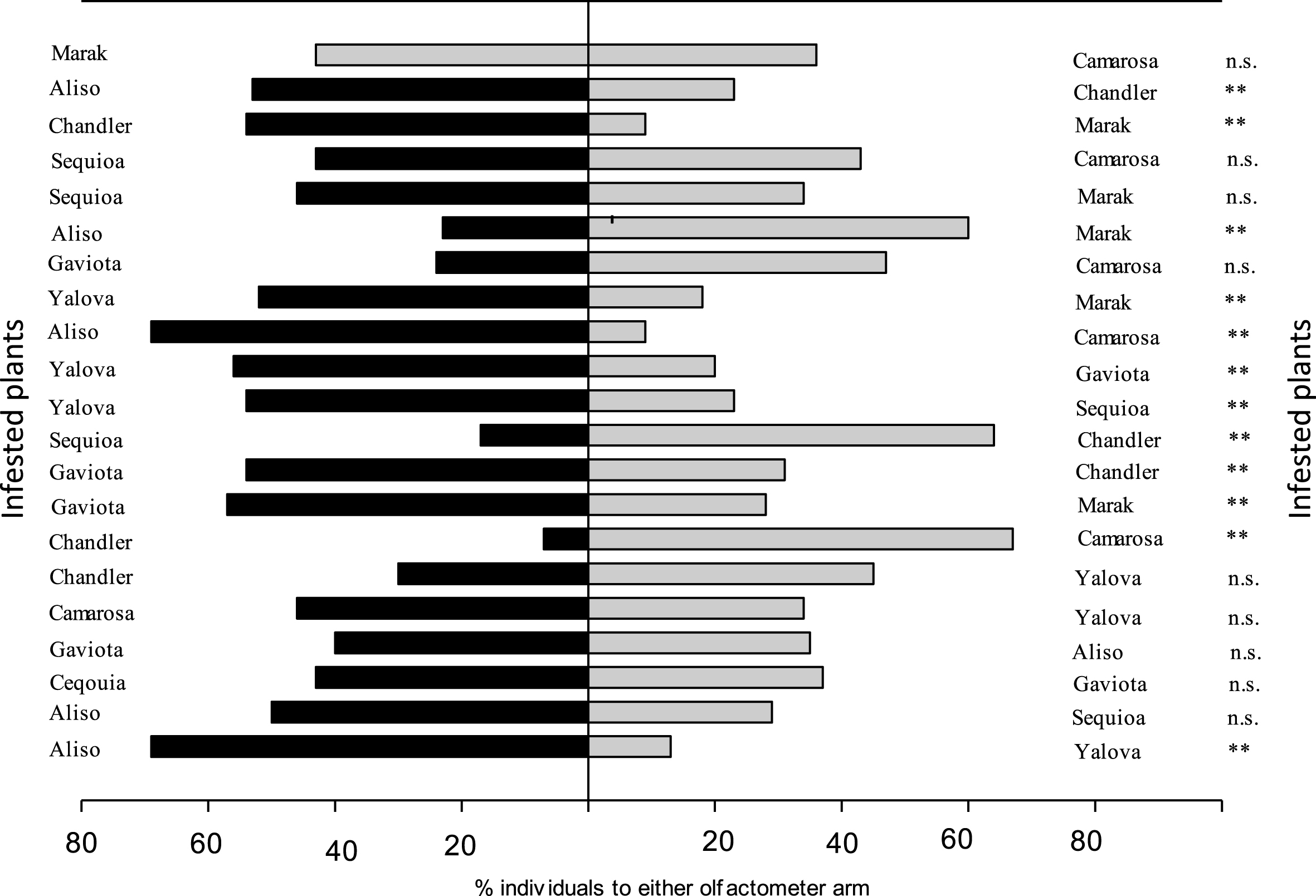
When presented with a choice between two strawberry cultivars among 21 tests, the predatory mite, N. californicus did not preferred odour emitted from each strawberry cultivar in nine dual tests, (Aliso & Gaviota), (Aliso & Seqouia), (Chandler & Gaviota), (Chandler & Yalova), (Gaviota & Seqoia), (Gaviota & Marak), (Gaviota & Camarosa), (Seqoia & Camarosa) and (Marak & Camarosa). The predator N. californicus preferred the odours emitted by some cultivars more than the others in another dual tests. Moreover, the odours of some of the strawberry cultivars, infested by T. urticae evoked a greater behavioral response than the other cultivars. The predatory mite significantly preferred spider mite- infested strawberry cultivar “Aliso “ and “ Chandler” over than the other cultivars. These cultivars were the most attractive for the predatory mite (Table 4 & Fig. 4).
Table 4
Response of Neoseiulus californicus to odours of different strawberry cultivars infested with Tetranychus urticae in a laboratory two-choice test
| Source of odour | Gh | df | Pvalue | Gp | df | Pvalue | GT | df | Pvalue |
| ‘Aliso’ with T. urticae vs. ‘Chandler’ with T. urticae | 1.57 | 2 | 0.45 | 5.94 | 1 | 0.014** | 7.51 | 3 | 0.06 |
| ‘Aliso’ with T. urticae vs. ‘Gaviota’ with T. urticae | 0.80 | 2 | 0.67 | 0.13 | 1 | 0.71n . s . | 0.93 | 3 | 0.81 |
| ‘Aliso’ with T. urticae vs. ‘Ceqouia’ with T. urticae | 5.27 | 2 | 0.07 | 2.16 | 1 | 0.14n . s . | 7.43 | 3 | 0.06 |
| ‘Aliso’ with T. urticae vs. ‘Yalova’ with T. urticae | 1.26 | 2 | 0.53 | 16.62 | 1 | 0.0004** | 17.88 | 3 | 0.0005 |
| ‘Aliso’ with T. urticae vs. ‘Camarosa’ with T. urticae | 0.027 | 2 | 0.98 | 18.59 | 1 | 0.0002** | 18.62 | 3 | 0.0003 |
| ‘Aliso’ with T. urticae vs. ‘Marak’ with T. urticae | 5.29 | 2 | 0.07 | 6.04 | 1 | 0.01** | 11.33 | 3 | 0.01 |
| ‘Chandler’ with T. urticae vs. ‘Gaviota’ with T. urticae | 4.13 | 2 | 0.13 | 2.16 | 1 | 0.14n . s . | 6.29 | 3 | 0.098 |
| ‘Chandler’ with T. urticae vs.’ Ceqoia’ with T. urticae | 1.06 | 2 | 0.59 | 10.63 | 1 | 0.0001** | 11.70 | 3 | 0.008 |
| ‘Chandler’ with T. urticae vs. ‘Yalova’ with T. urticae | 2.16 | 2 | 0.34 | 1.21 | 1 | 0.27n . s . | 3.37 | 3 | 0.34 |
| ‘Chandler’ with T. urticae vs. ‘Marak’ with T. urticae | 0.21 | 2 | 0.90 | 12.97 | 1 | 0.0003** | 13.19 | 3 | 0.004 |
| ‘Chandler’ with T. urticae vs. ‘Camarosa’ with T. urticae | 0.07 | 2 | 0.96 | 25.64 | 1 | 0.0000** | 25.71 | 3 | 0.0001 |
| ‘Gaviota’ with T. urticae vs. ‘Cequioa’ with T. urticae | 0.36 | 2 | 0.83 | 0.14 | 1 | 0.70n . s . | 0.50 | 3 | 0.92 |
| ‘Gaviota’ with T. urticae vs. ‘Yalova’ with T. urticae | 0.21 | 2 | 0.90 | 7.85 | 1 | 0.005** | 8.04 | 3 | 0.04 |
| ‘Gaviota’ with T. urticae vs. ‘Marak’ with T. urticae | 0.30 | 2 | 0.86 | 3.40 | 1 | 0.06n . s . | 3.69 | 3 | 0.30 |
| ‘Gaviota’ with T. urticae vs. ‘Camarosa’ with T. urticae | 0.029 | 2 | 0.98 | 3.18 | 1 | 0.07n . s . | 3.21 | 3 | 0.36 |
| ‘Cequioa’ with T. urticae vs. ‘Yalova’ with T. urticae | 0.37 | 2 | 0.83 | 4.61 | 1 | 0.03* | 4.98 | 3 | 0.17 |
| ‘Cequioa’ with T. urticae vs. ‘Marak’ with T. urticae | 0.05 | 2 | 0.97 | 0.57 | 1 | 0.45n . s . | 0.62 | 3 | 0.89 |
| ‘Cequioa’ with T. urticae vs. ‘Camarosa’ with T. urticae | 0.80 | 2 | 0.67 | 0.56 | 1 | 0.44n . s . | 0.80 | 3 | 0.85 |
| ‘Yalova’ with T. urticae vs. ‘Marak’ with T. urticae | 1.50 | 2 | 0.41 | 7.57 | 1 | 0.005n . s . | 90.7 | 3 | 0.028 |
| ‘Yalova’ with T. urticae vs. ‘Marak’ with T. urticae | 9.14 | 2 | 0.047 | 0.57 | 1 | 0.45n . s . | 6.71 | 3 | 0.082 |
| ‘Marak’ with T. urticae vs. ‘Camarosa’ with T. urticae | 1.47 | 2 | 0.48 | 0.29 | 1 | 0.59n . s . | 1.76 | 3 | 0.62 |
*: P < 0.05, **: P < 0.01, n . s .: not significant.
Fig.4
Responses of adult female of Neoseiulus californicus in Y-tube olfactometer when offer infested seven strawberry cultivars leaves. G test was used to evaluate whether the result differed from a 50 : 50 distribution between the two olfactometer arms (*P < 0.05, **P < 0.01, n.s. not significant).
4Discussion
Neoseiulus californicus is an effective biological control agent against T. uticae and other spider mite pests of agricultural crops [31, 32]. This predator can be used for controlling T. urticae on strawberry [33]. The predatory mite showed a significant preference for the volatiles emitted from uninfested strawberry plants. In this study, the predatory mite attracted the uninfested plant over clean air. Several studies confirmed that uninfested plants are attractive for the phytoseiid mites [29, 34–36]. Zhang et al. (2011) [37] were reported that N. californicus preferred the odours of undamaged plants such as eggplant, pepper and tomato to clean air. This might be due to previous experience of the predators. In this study, the predatory mite was reared on strawberry leaves for at least one generation and previous experience of the predator affect on the response of the predator. Uninfested plant of some strawberry cultivars such as ‘Sequioa’ was not attractive for the predatory mite. It might be due to the volatiles of this cultivar which were not preferred by the predator. Takabayashi & Dicke [28] were reported about the effect of previous experiences on the response of phytoseiid mites to volatiles emitted from uninfested plants. Other studies indicated that the phytoseiid mites prefer uninfested plants over clean air, e.g. Neoseiulus cucumeris (Oudemans), showed a significant preference for volatiles from infested cucumber leaves with or without Thrips tabaci (Lindeman) over clean air [38].
The predatory mite N. californicus preferred infested plants when given a choice between T. urticae-infested strawberry cultivars and clean air. Several studies have reported that N. californicus discriminate between plants infested with spider mite and uninfested plants over clean air [39, 40]. Neoseiulus californicus is attracted to volatiles emitted from T. urticae-infested lima bean leaves, volatiles emitted from artificially damaged lima bean leaves, and volatiles emitted from feces, exuviate and eggs of T. urticae. Moreover, N. californicus is attracted to synthetic compounds such as (Z)-3-hexen-1-ol, (E)-2-hexenal and (Z)-3-hexenyl acetate, which are commonly released from various plants even when artificially damaged [39]. Shimoda (2010) [41] showed that methyl salicylate and linalool are key volatiles that elicit strong olfactory responses in N. californicus. The volatiles emitted from plants infested with T. urticae attract predators, Other researchers investigated that, e.g. Sabelis & van den Boam (1983) and van Wijk et al. (2008) [14, 34] showed that Phytoseiulus persimilis Athias- Henriot preferred infested bean leaves over clean air. Neoseiulus barkeri (Athias-Henriot) preferred infested plants over clean air. The predatory mite’s ability to find prey-infested plants is important trait to be considered when selecting a predator for biological control purpose [3]. Phytoseiulus macropilis (Banks) were able to use odours from T. urticae infested strawberry plant to locate prey in olfactometer experience [42].
Comparing the response of N. californicus to infested vs. uninfested plants showed that the predators prefer infested plants. Some researchers reported similar results indicating that phytoseiid mites prefer T.urticae-infested plants over uninfested plants [6, 26, 43]. Neoseiulus cucumeris can discriminate between odours emitted from tulip bulbs infested with rust mites (Aceria tulipae (Keifer)), odours from uninfested bulbs and artificially wounded bulbs [44]. Phytoseiulus persimilis preferred odours emitted from infested cucumber plants over odours emitted from uninfested cucumber plants [45]. Neoseiulus womersleyi (Schicha) were attracted to volatiles emitted from the tea plant infested with T. kanazawai Kishida more than uninfested tea leaves [46]. Typhlodromus kerkirae Swirski & Ragusa is attracted to T. urticae-infested leaves more than non-infested bean leaves [47]. Neoseiulus cucumeris (oudemans) can discriminate between odours emitted from bulbs infested with A. tulipae and odours emitted from uninfested bulbs [44].
Maeda et al. (2000) [46] demonstrated that rearing condition is an important factor in the olfactory response of a predatory mite. The feeding history affects the response of a predatory mite to volatiles emitted from plants infested with their prey. For example, T. kerkirae are attracted to volatiles emitted from plants infested with spider mites [15]; or N. womersleyi is attracted only to volatiles of the plants on which they had previous experience [18]. Drukker et al. (2000) [17] showed that If a predatory mite is reared in the absence of plant odours, it will not be attracted to the odours of the plants that are infested with its prey. Krips et al. (1999) [26] showed that P. persimilis which were reared on lima bean plants infested with spider mite were not attracted to spider mite-induced volatiles from gerbera. In contrast, the predator that was reared on gerbera leaves infested with spider mite was strongly attracted to volatiles from spider mite-infested gerbera. The response to spider mite-damaged cucumber leaves increased significantly after the predators were given several days of experience with spider mites on cucumber [3, 6, 28]. In our study, the predatory mite N. californicus prefers the odour of the spider mites on strawberry plants may be the predators were reared on strawberry leaves that were infested with T. urticae.
The efficacy of biological control of T. urticae by releasing a general predator such as N. californicus is influenced by plant species. Several species of host plants respond to damage of spider mites through the production of volatiles that attract P. persimilis. Predators locate prey habitat by using chemical cues emitted by plants. The predatory mites are capable of exploiting herbivore-induced volatiles emitted from the prey-infested plants as a foraging cue [38, 49]. Takabayashi & Dicke (1992) [28] found that host plants on which predatory mites consume their prey affect attraction of the predators to the plant volatiles. Zhong et al. (2011) [37] studied the olfactory response of N. cucumeris to odours from eggplant, pepper and tomato. The response of P. persimilis varies with host-plant species [6, 33, 28]. Tatemoto & Shimoda (2008) [38] investigated the response of N. cucumeris to volatiles associated with two different plant species (cucumber and onion) infested with T. tabaci.
In conlcusion, N. californicus was attracted to uninfested and infested strawberry plants, indicating that biological conotrl of the spider mites can be conducted optimisticlly. Among tested strawberry cultivars, ‘Aliso’ and ‘Chandler’ were more attractive for the predatory mite, indicating that there might be special volatiles emitted from these cultivars that elicit a strong olfactory response in N. californicus. It is recommended that further investigation in to these cultivars. Further investigation the olfactory responses to synthetic volatiles and theoretical studies with mathematical models should be conducted.
References
[1] | Dicke M . Evolution of induced indirect defence of plants. In: Tollrian R, Harvell CD. (Eds.). Evolution of Inducible Defenses. Princeton University Press. Princeton. New Jersey. (1999) ; pp. 483–520. |
[2] | Dicke M , Sabelis MW , Takabayashi J , Bruin J , Posthumus MA . Plant strategies of manipulating predator prey interactions through allelochemicals: Prospects for application in pest control. Journal of Chemical Ecology. (1990) ;16: (11):3091–118. |
[3] | Turlings TCJ , Wackers FLL , Vet EM , Lewis WJ , Tumlinson JH . Learning of host finding cues by hymenopterous parasitoid. In: Papaj DR, Lewis AC, (eds.). Insect Learning. (1993) ; pp. 51–78. |
[4] | Vet LEM , Dicke M . Ecology of infochemical use by natural enemies in tritrophic context. Annual Review of Entomology. (1992) ;37: :141–72. |
[5] | Van den Boom CE , Van Beek TA , De Groot A , Dicke M . Attraction of Phytoseiulus persimilis (Acari: Phytoseiidae) towarda volatiles from various Tetranychus urticae- infested plant species. Bulletin Entomological Research. (2004) ;92: :539–46. |
[6] | Takabayashi J , Dicke M , Takahashi S , Posthumud MA , van Beek TA . Leafage affects the composition of herbivore- induced summons and attraction of predatory mites. Journal of Chemical Ecology. (1994) ;20: :373–86. |
[7] | Takabayashi J , Takahashi S , Dicke M , Posthhumus MA . Development stage of herbivore Pseudaletia separate affects production of herbivore- induced synomone by corn plants. Journal of Chemical Ecology. (1995) ;21: :273–87. |
[8] | Nachappa P , Margolies DC , Nechols JR , Loughin T . Phytoseiulus persimilis response to herbivore-induced plant volatiles as a function of mite-days. Experimental & Applied Acarology. (2006) ;40: (3-4):231–9. |
[9] | Dicke M , Takabayashi J , Posthumus MA , Schutte C , Olga EK . Plant-phytoseiid interactions mediated by herbivore-induced plant volatiles: Variation in production of cues and in responses of predatory mites. Experimental and Applied Acarology. (1998) ;22: :311–33. |
[10] | Takabayashi J , Dicke M , Posthumus MA . Variation in composition of predator attracting allelochemicals emitted by herbivore-infested plant: Relative influence of plant and herbivore. Chemoecology. (1991) ;2: :1–6. |
[11] | Sabelis MW , Van de Baan HE . Location of distant spider mite colonies by phytoseiid predatores: Demonstration of specific kairomones emitted by Teranychus urticae and Panonychus ulmi. Entomologia Experimentalis et Applicata. (1988) ;33: :303–14. |
[12] | De Moraes CM , Lewis WJ , Pare PW , Alborn HT , Tumlinson JH . Herbivore-infested plants selectively attract parasitoids. Nature. (1998) ;393: :570–73. |
[13] | De Boer JG , Posthumus MA , Dicke M . Identification of volatiles that are used in discrimination between plants infested with prey or non-prey herbivores by a predatory mite. Journal of Chemical Ecology. (2004) ;30: :2215–29. |
[14] | van Wijk M , De Bruijn JA , Sabelis MW . Predatory mite attraction to herbivore- induced plant odours is not a conseguence of attraction to individual herbivore – induced plant volatiles. Journal of Chemical Ecology. (2008) ;34: :791–803. |
[15] | Koveos DS , Broufas GD . Feeding history affects the response of the predatory mite Typhlodromus kerkirae (Acari: Phytoseiidae) to volatiles of plants infested with spider mites. Experimental and Applied Acarology. (1999) ;23: :429–36. |
[16] | Maeda T , Liu Y , Ishiwari H , Shimoda T . Conditioned olfactory responses of a predatory mite, Neoseiulus womersleyi, to volatiles from prey-infested plants. Entomologia Experimentalis et Applicata. (2006) ;121: :167–75. |
[17] | Drukker B , Bruin J , Jacobs G , Kroon A , Sabelis MW . How predatory learns to cope with variability in volatile plant signals in the environment of their herbivorous prey. Experimental and Applied Acarology. (2000) ;24: :881–95. |
[18] | Maeda T , Takabayashi J , Yano S , Takafuji A . The effects of rearing conditions on the olfactory response of predatory mites, Phytoseiulus persimilis and Amblyseius womersleyi (Acari: Phytoseiidae). Applied Entomology and Zoology. (2000) ;35: :345–51. |
[19] | Ishiwari H , Suzuki T , Maeda T . Essential compound in herbivore- induced plant volatiles that attract the predatory mites Neoseiulus womersleyi. Journal of Chemical Ecology. (2007) ;33: :1670–81. |
[20] | Helle W , Sabelis MW . Spider mites: Their biology, natural enemies and control. World crop pests, vol 1A: . (1985) ; Elsevier, Amsterdam. |
[21] | Greco NM , Sanchez NE , Liljesthrom GG . Neoseiulus californicus (Acari: Phytoseiidae) as a potential control agent of Tetranychus urticae (Acari: Tetranychidae): Effect of pest/predator ratio on pest abundance on strawberry. Experimental & Applied Acarology. (2005) ;37: :57–66. |
[22] | Gerson U , Weintraub PG . Mites for the control of pests in protected cultivation. Pest Managment Science. (2007) ;63: :658–76. |
[23] | Sato ME , Silva MZ , De Souuza Filho MF , Matioli AL , Raga A . Management of Tetranychus urticae (Acari: Tetranychidae) in strawberry field with Neoseiulus californicus (Acari: Phytoseiidae) and acaricids. Experimental & Applied Acarology. (2007) ;42: :107–20. |
[24] | Fraulo AB , Liburd OE . Biological control of two spotted spider mite, Tetranychus urticae, with predatory mite, Neoseiulus californicus, in strawberries. Experimental & Applied Acarology. (2007) ;43: :109–19. |
[25] | Dicke M , Sabelis MW , Jong M . Analysis of prey preference in phytoseiid mites by using an olfactometer, predation models and electrophoresis. Experimental and Applied Acarology Acarology. (1988) ;5: :225–41. |
[26] | Krips OE , Kleijn PW , Willems PEL , Gols GIZ , Dicke M . Leaf hairs influence searching efficiency and predation rate of the predatory mite Phytoseiulus persimilis (Acari: Phytoseiidae). Experimental & Applied Acarology. (1999) ;23: :119–31. |
[27] | Himanen S , Vuorinen T , Tuovinen T , Holopainen JK . Effect of cyclamen mite (Phytosemus pallidus) and leaf beetle (Galerucella tenella) damage on volatile emission from strawberry plants and orientation of predatory mite (Neoseiulus cucumeris, N. californicus, Euseius finlandicus). Journal of Agriculture Food Chemistry. (2005) ;53: :8624–30. |
[28] | Takabayashi J , Dicke M . Response of predatory mites with different rearing histories to volatiles of an infested plant. Entomologia Experimentalis et Applicata. (1992) ;64: :187–93. |
[29] | Sabelis MW , van derWeel JJ . Anemotactic responses of the predatory mite, Phytoseiulus persimilis Athias-Henriot, and their role in prey finding. Experimental and Applied Acarology. (1993) ;17: :1–9. |
[30] | Sokal R , Rohlf FJ . Biometry, 3rd ed., W. H. Freeman and Company, San Francisco, CA, (1995) . |
[31] | Garcia- Mari F , Gonzalez–Zamora JE . Biological control of Tetranychus urticae (Acari: Experimental and Applied Acarology. (1999) ;23: :487–95. |
[32] | Easterbrook MA , Fitzgerald JD , Solomon MG . Biological control of strawberry tarsonemid mite Phytonemus pallidus and two-spotted spider mite Tetranychus urticae on strawberry in the UK using species of Neoseiulus (Amblyseius) (Acari: Phytoseiidae). Experimental and Applied Acarology. (2001) ;25: :25–36. |
[33] | Greco NM , Liljesthrom GG , Sanchez NE . Spatial distribution and coincidence of Neoseiulus californicus and Tetranychus urticae (Acari: Phytoseiidae, Tetranychidae) on strawberry. Experimental & Applied Acarology. (1999) ;23: :567–80. |
[34] | Dicke M , Sabelis MW . How do plants obtain predatory mites as bodyguards? Netherlands Journal of Zoology. (1988) ;38: :148–65. |
[35] | De Boer JG , Dicke M . Experience with methyl salicylate affects behavioral response of a predatory mite to blends of herbivore- induced plant volatiles. Entomologia Experimentalis et Applicata. (2004) ;110: :181–9. |
[36] | Maleknia B , Glopaiganie AZ , Saboori A , Magalhas S . Olfactory response of Phytoseiulus persimilis to rose plant with or without prey or competitors. Acarologia. (2013) ;53: :235–46. |
[37] | Zhong F , Rong YH , Gao Y , Qi G , Zhao CY , Lu LH . Erratum to: Olfactory responses of Neoseiulus cucumeris (Acari: Phytoseiidae) to odours of host plants and Frankliniella occidentalis (Thysanoptera: Thripidae) plant complexes. Arthropod-Plant Interactions. (2011) ;5: :315–7. |
[38] | Tatemoto S , Shimoda T . Olfactory responses of the predatory mites (Neoseiulus cucumeris) and insects (Orius strigicollis) to two different plant species infested with onion thrips (Thrips tabaci). Journal of Chemical Ecology. (2008) ;34: :605–13. |
[39] | Shimoda T , Ozawa R , Sano K , Yano E , Takabayashi J . The involvement of volatile infochemicals from spider mites and from food-plants in prey location of the generalist predatory mite Neoseiulus californicus. Journal of Chemical Ecology. (2005) ;31: :2019–32. |
[40] | Emami K , Golpayegani AZ , Saboori A . Article Diet dependent olfactory response and predation rate of Neoseiulus californicus (Acari: Phytoseiidae) in the presence of Frankliniella occidentalis and Tetranychus urticae, Persian Journal of Acarology. (2015) ;4: (1):95–109. |
[41] | Shimoda T . A key volatile infochemical that elicits a strong olfactory response of the predatory mite Neoseiulus californicus, an important natural enemy of the two-spotted spider mite Tetranychus urticae. Experimental & Applied Acarology. (2010) ;50: (1):9–22. |
[42] | Oliveria H , Fadini MAM , Venzon M , Rezende M , Rezende F , Pallini A . Evaluation of the predatory mite Phytoseiulus macropilis (Acari: Phytoseiidae) as a biological control agent of the two spotted spider mite on strawberry plant under greenhouse condition. Experimental & Applied Acarology. (2009) ;47: :275–83. |
[43] | Bruin J , Dicke M , Sabelis MW . Plants are better protected against spider-mites after exposure to volatiles from infested conspecifics. Experientia. (1992) ;48: :525–9. |
[44] | Aratchige NS , Lesna I , Sabelis MW . Below-ground plant parts emit herbivore-induced volatiles: Olfactory responses of a predatory mite to tulip bulbs infested by rust mites. Experimental and Applied Acarology. (2004) ;33: :21–30. |
[45] | Janssen A . Plant with spider mites prey attract more predatory mites than clean plant under greenhouse condition. Entomologia Experimentalis et Applicata. (1999) ;90: :191–8. |
[46] | Maeda T , Liu Y . Intraspecific variation in the olfactory response of the predatory mite Neoseiulus womersleyi Schicha (Acari: Phytoseiidae) to different amount of spider mite-infested plant volatiles. Applied Entomology and Zoology. (2006) ;41: (2):209–15. |
[47] | Dimitri SK , George DB . Feeding history affects the response of the predatory mite Typhlodromus kerkirae (Acari: Phytoseiidae) to volatiles of plants infested with spider mites. Experimental Applied Acarology. (1999) ;23: :429–36. |
[48] | Sabelis MW , Afman BP . Synomone-induced suppression of take-off in the phytoseiid mite Phytoseiulus persimilis Athias-Henriot. Experimental and Applied Acarology. (1994) ;18: :711–21. |
[49] | Satoshi T , Takeshi S . Olfactory response of the predatory mite (Neoseiulus cucumeris) and insect (Orius strigicollis) to two different plant species infested with onion thrips (Thrips tabaci). Journal of Chemistry Ecology. (2008) ;34: :605–13. |

